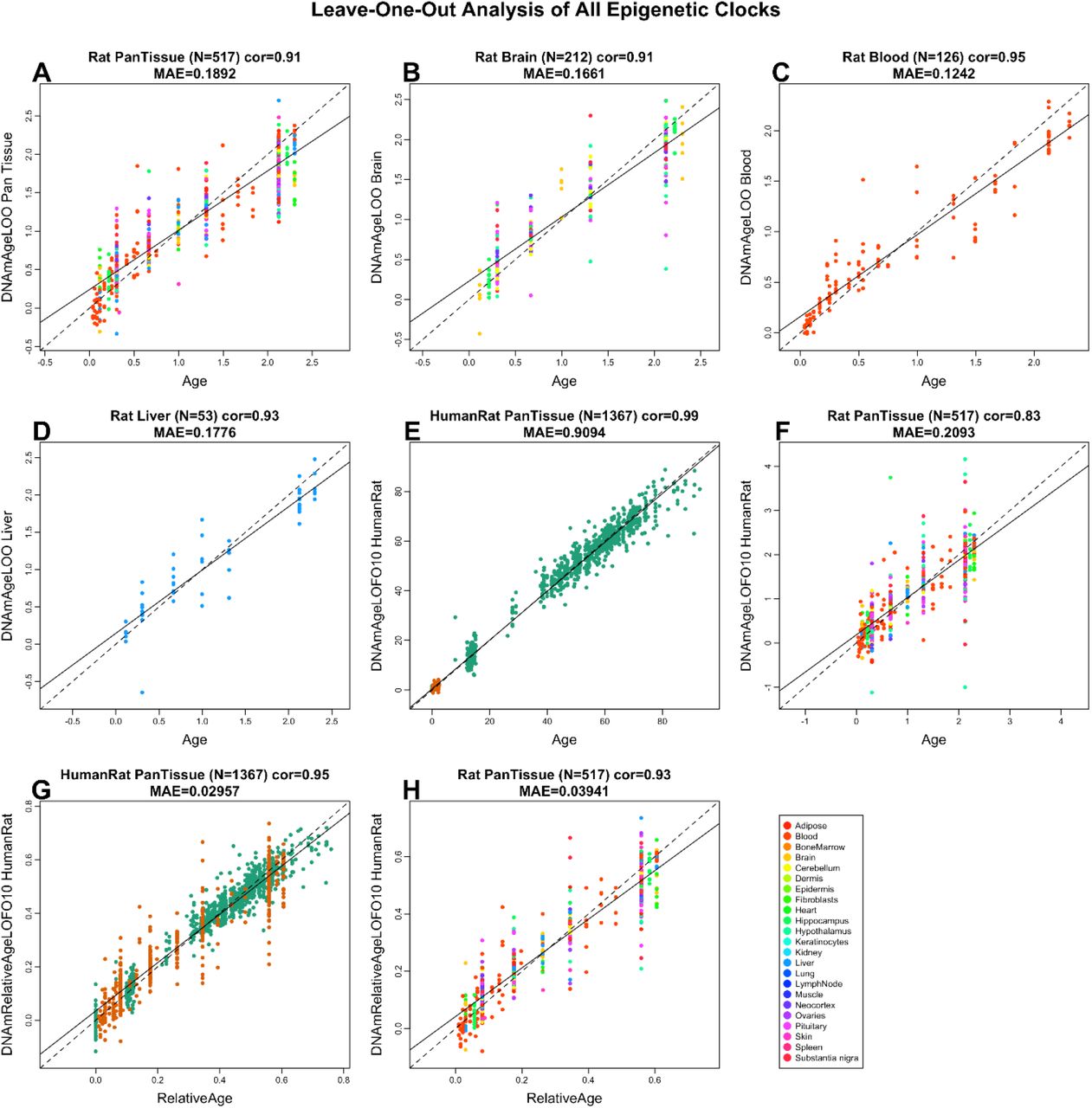
Данная статья собрана из нескольких моих ранних заметок о влиянии внеклеточного матрикса на процесс старения. Текст статьи будет обновляться — я планирую дополнить список литературы и внести некоторые стилистические коррективы. Denis Odinokov
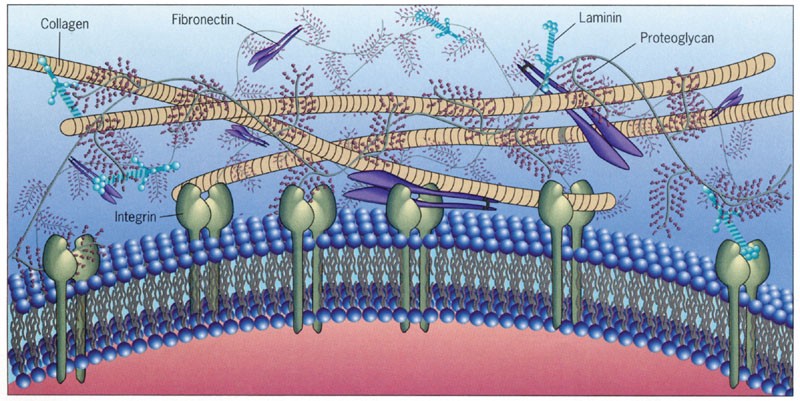
Соединительная ткань включает в себя волокнистые ткани, жировую ткань, хрящи, кости, костный мозг, кровь и включает значительное количество внеклеточных веществ, которые образуют структуру органов и тканей организма. Разнообразные типы коллагенов являются основным компонентом соединительной ткани позвоночных и составляют примерно одну треть белков в организме.
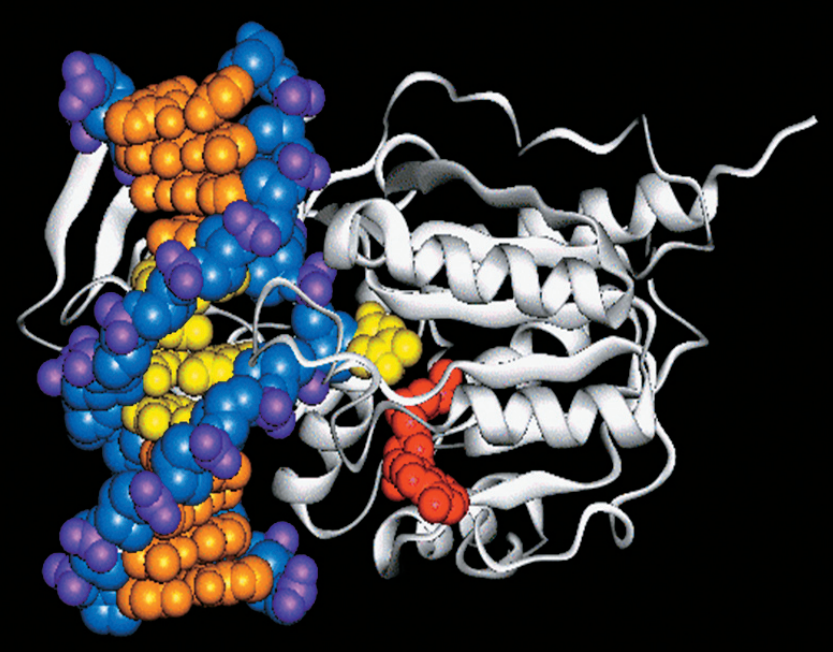
Влияние образования поперечных сшивок между фибрилами коллагена— превалирующего белка внеклеточного матрикса — на процесс старения и связанные со старением заболевания изучалось в ранних работах Йохана Бьоркштена (Johan Björksten), которые датируются 1942 годом. Автор предположил, что образование внутри- и межмолекулярных ковалентных сшивок меняет структуру макромолекул и приводит к их дисфункции: «Старение живых организмов, на мой взгляд, связано с случайным образованием […] мостов между молекулами белка, которые не могут быть разрушены клеточными ферментами» [2].
Фриц Верзар (Fritz Verzár) впервые продемонстрировал в 1950-х годах, что количество «сшитых» коллагеновых волокон внеклеточного матрикса экспоненциально увеличивается с возрастом [3].
Эти модифицированные белки остаются невосстановленными, накапливаются на протяжении всей жизни и вызывают изменения в механических свойствах микроокружения, тканей и органов, что приводит к порочному циклу постепенного увеличения повреждений, и серьезно ограничивают применение сенолитиков, стволовых клеток и других методов «антивозрастной» терапии.
Ни один новомодный метод, обсуждаемый в связи с влиянием на продолжительность жизни, как-то экзосомы стволовых клеток, донорские митохондрии, сенолитики, эпигенетическое манипуляции (список можно продолжить) — не способен восстановить структуру матрикса, физические свойства которого (структура и жёсткость, в основном) являются не менее сильным, чем химический (то есть через ионы и молекулы) сигнальным фактором, определяющим судьбу клеток, тканей, органов и организма в целом.
Кроме интегрирующей роли и связи матрикса с вегетативной нервной системой, которая ранее была изучена Альфредом Пишингер и Хартмут Хайне (Alfred Pischinger, Hartmut Heine) [4], следует отметить что молекулы коллагена содержат большое количество пролина — аминокислоты способной проводить слабые электромагнитные волны, генерируемые клетками и тканями. Поэтому нельзя исключить возможность, что соединительные ткани, в дополнение к механическим и химическим сигналам, формируют единую биоэлектрическую сигнальную систему организма. Тут интересно отметить ряд работ, в которых исследователям удалось, меняя топографию среды [5, 6] или электромагнитное излучение [7, 8], не только управлять клеточным циклом, но и превращать взрослые соматические клетки в стволовые без помощи вирусов с вектором факторов Яманака.
Даже незначительные изменения геометрии межклеточной среды оказывают влияние на экспрессию генов и функцию клеток. Передаваемые через цитоскелет клетки механические сигналы и жесткость ядерной ламины (ядерной пластинки) совместно регулируют динамику ядра и хроматина [9, 10].
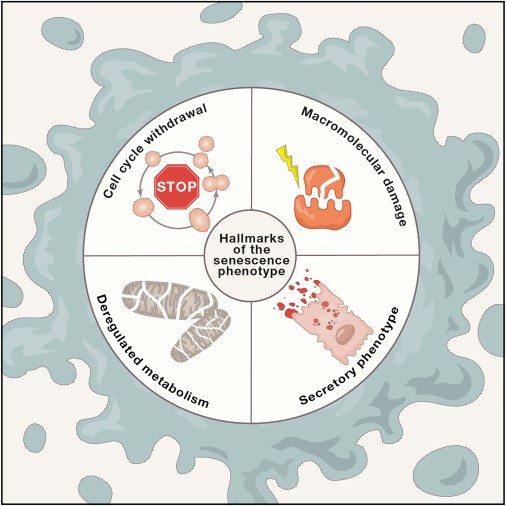
Так, «молодые» фибробласты стареют в старом матриксе и наоборот — «старые» клетки утрачивают признаки связанного со старением секреторного фенотипа в «молодом» матриксе [11–13].
Вероятно, что жесткость внеклеточного матрикса также влияет на морфологию митохондрий и синтез АТФ [14, 15].
Некоторые могут возразить «давайте разрушим старый матрикс — ведь как-то организм обновляет коллаген — и создадим новый, лучше прежнего» игнорируя тот факт, что, во-первых, молекулы коллагена кожи, согласно ряду исследований, имеют период полужизни 15 лет, а хрящей — более 100 лет [16–18], и во-вторых, на сегодняшний день не существует избирательных ферментов и разрушать придется все протеины. Более того, по мере увеличения количества сшивок, коллагеновые фибриллы становятся более плотными, делая матрикс менее доступным для ферментов, участвующих в естественном обороте коллагенов. Следовательно, скорость оборота белков матрикса со временем замедляется, и как результат — это приводит к появлению еще большего количества сшивок.
Процесс неферментатиивного гликозилирования или гликирования — причины образования сшивок — практически не регулируется. Хотя есть возможность «сдерживания» гликозилирования через трансгликацию [19], при которой в «расход» идут глутатион, полиамины, тиолы, свободные аминокислоты, например, таурин, лизин, предотвращая формирование новых сшивок.
Описание прочих негативных эффектов гликирования, отличных от изменения структуры и жесткости внеклеточного матрикса, например, формирование метаболитической памяти [20], увеличение воспалительных процессов и т.п., выходит за пределы данной статьи.
Также есть варианты уменьшения вреда ответной реакции организма на увеличение жесткости внеклеточного матрикса — секрецию трансформирующего ростового фактора бета (TGF-beta), богатого цистеином белка 61 (Cyr61/CCN1) и прочих сигнальных молекул. Например, фиброз тканей может быть замедлен ингибиторами матриксных металлопротеиназ, одним из которых является доксициклин.
Ранее разработанные препараты против поперечных сшивок, таких как Alagebrium/ALT-711 [21], C36 [22], TRC4149 [23] были нацелены на нейтрализацию карбоксиметиллизина — самого распространённого позднего продукта реакции Майяра, накапливающемся в организме при диабете.
Считается, что глюкезепан, оказывает наибольшее влияние на течение болезней пожилого возраста человека и поэтому является приоритетной мишенью для противовозрастной терапии.
Однако, даже появление обещанного Дэвидом Шпигель решения — его группа работает над созданием ферментов против глюкозепана [24], не решит проблемы — разрушение одной из десятков видов известных сшивок, скорее всего, будет иметь временный характер и не сможет значительно изменить физические свойства межклеточного матрикса.
Кроме образования поперечных сшивок, белки также подвержены рацемизации [25–32].
Рацемизация — это спонтанный процесс, в результате которого происходит превращение оптически активных соединения в рацемическую смесь. Так, аминокислоты превращаются из L-формы в зеркальную D-форму (часть из них имеют свою биологическую активность). Рацемизация происходит во всех белках, но протеолитические ферменты не дают им накапливаться. Однако, в тканях с замедленным метаболизмом, количество рацемированных белков линейно увеличивается, например, в тканях зубов на 0,1% в год.
Рацемизация — это процесс «естественного» старения белков, поэтому может использоваться в качестве молекулярного индикатора старения, а также для идентификации долгоживущих белков. Количественное измерение степени рацемизации аспарагиновой кислоты используется в криминалистике и судебно-медицинской экспертизе для определения возраста останков.
Шведский химик Вернер Кун (Werner Kuhn) в 1955 году предположил, что данные изменения оптически активных соединений вызывают старение организма [33, 34]. Действительно, накопление белков с аномальными физико-химическими свойствами способствуют прогрессу ряда состояний, связанных со старением, таких как атеросклероз, эмфизема легких, пресбиопия, катаракта, дегенеративные заболевания хрящей и возрастные заболевания нервной системы.
Потеря функций белков происходит из-за протеолиза «поломанных» белков или из-за изменения их молекулярной структуры.
Поперечные сшивки коллагенов внеклеточного матрикса затрудняют доступ протеолитических и исправляющих ферментов (l-isoaspartyl methyltransferase [35]) к структурно измененным в результате рацемизации белкам [36]. Это способствует процессу накопления поврежденных белков в коллагене, что усугубляет ситуацию и негативно сказывается на механических свойствах тканей при старении [18].
Здесь я хочу поблагодарить Александра Фединцева за идею и дополнения дальнейшей части данного текста.
Итак, подвергшиеся неферментативному гликозилированию (гликированию), то есть реакции между углеводами (глюкозой, фруктозой и прочими) и свободными аминогруппами, белки, а также липиды и нуклеиновые кислоты, образуют так называемые конечные продукты гликозилирования (КПГ).
Роль КПГ в формировании внутри- и межмолекулярных поперечных сшивок белков хорошо изучена и не вызывает сомнений.
Кроме гликирования, существует не менее важный процесс, протекающий как в патологических, так и нормальных условиях — перекисное окисление липидов (ПОЛ).
Оба процесса, перекисное окисление липидов и гликирование, включают целую сеть различных реакций, в которых получается необычайно сложная смесь соединений [37].
Многочисленные исследования указывают на связь липидного состава мембран с максимальной продолжительностью жизни разных видов животных. Виды с преобладающим содержанием насыщенных жирных кислот в составе клеточных мембран обладают большей продолжительностью жизни даже по сравнению с филогенетически близкими видами [38–59].
Более того, эта разница в составах мембран позволяет объяснить «парадокс птиц»: птицы имеют очень быстрый метаболизм, но живут на порядок дольше млекопитающих со схожей величиной основного обменом (basal metabolic rate). Выяснилось, что у птиц индекс пероксидации клеточных мембран значительно ниже.
Также, липидный состав мембран позволяет объяснить разницу между продолжительностью жизни рабочих пчел и королев без привлечения пресловутых «программ старения».
Насыщенные и мононенасыщенные жирные кислоты более стабильны и менее подвержены перекисному окислению, по сравнению с полиненасыщенными жирными кислотами (ПНЖК). Это происходит потому, что ПНЖК имеют протоны в уязвимой бис-аллильной позиции. Такой протон легко оторвать («абстрагировать») от молекулы жирной кислоты и это является первым шагом в цепочке реакций перекисного окисления липидов.
Распространенное мнение о том, что ПНЖК безусловно полезны стоит подвергнуть критике. Хотя возможно, что ПНЖК полезны как раз тем, что создают умеренный оксидативный стресс, способствуя гормезисному стресс-ответу.
Ряд исследований указывает на количественную связь между процессом перекисного окисления липидов и образованием поперечных сшивок белков матрикса. Так, например, один из продуктов перекисного окисления липидов, малондиальдегид (возникает при деградации ПНЖК), образует такое же количество сшивок с белками, как и глюкоза [60].
Это позволяет предположить, что долгоживущие виды не только в меньшей степени страдают от ПОЛ, но и имеют более замедленный, за счет уменьшения реакционной способности жирных кислот, процесс изменения белков внеклеточного матрикса.
Более десяти лет назад российский ученый Михаил Щепинов предложил использовать жирные кислоты, у которых водород заменен на дейтерий (изотоп, имеющий больший атомный вес и более прочную связь с атомом углерода), для продления жизни и лечения ряда заболеваний, вызванных избыточным синтезом свободных радикалов [61–64]. Измененные жирные кислоты более устойчивы к окислению и предотвращают разрушение клеточной мембраны.
В настоящее время, Retrotope — компания Михаила Щепилова, ожидает подтверждение FDA на проведение последней стадии клинических испытаний экспериментального препарата RT001. Недавно, dPUFA продемонстрировал задержку развития заболевания у 2 пациентов с болезнью Зейтельбергера, врожденной патологией характеризующееся нарушением обмена веществ с прогрессирующей липоидной дегенерацией в центральной нервной системе.
Возможно, что это лекарство поможет не только больным с наследственной нейродегенерацией, но также сможет значительно сдерживать старение за счет уменьшения перекисного окисления липидов и, как следствие, замедления процесса образования поперечных сшивок коллагенов внеклеточного матрикса.
Все обновления, а также ссылки на литературу смотрите в оригинале статьи.
27.10.2018 Источник: medium.com