Введение
Гетерохронный парабиоз - это процедура, при которой молодое животное и старое животное хирургически соединяют так, чтобы через определенное время у двух партнеров была общая кровеносная система [1,2,3]. Исследования парабиоза дали множество открытий относительно механизмов, лежащих в основе старения ниш стволовых клеток. Было показано, что у старых партнеров улучшается здоровье во многих тканях, когда они делили кровь с более молодым животным [1, 4, 5]. Известная интерпретация гетерохронного парабиоза состоит в том, что старение податливо и что процесс старения может быть замедлен или даже обращен вспять [1, 6]. Таким образом, исследования гетерохронного парабиоза положили начало изучению системного старения и омоложения.
Старение мозга, в частности, связано с прогрессирующей потерей функциональности и, как полагают, в значительной степени является результатом чрезмерной активации микроглии, миелоидных клеток, находящихся в головном мозге [7]. Возрастное снижение функции мозга и когниции (познавание) (среди многих других функций организма) когда-то считалось неизбежным и постоянным [8]. Интересно, что исследования парабиоза поставили под сомнение это представление, продемонстрировав пластичность поддержания и функционирования мозга после изменения возраста крови, а также предоставления обогащенной окружающей среды и обобществленных молодых органов старым партнерам [5, 9, 10].
Было высказано предположение, что несколько системных белков и инфузия молодой плазмы влияют на пластичность старения мозга [9, 11,12,13,14,15,16,17], хотя и с некоторыми противоречиями в отношении фактических возрастных уровней некоторых из этих факторов кандидатов, таких как GDF11, B2M, CCL11 и TIMP2 [9, 18,19,20,21,22,23,24]. В исследованиях инфузии плазмы молодых людей также не наблюдалось увеличения продолжительности здоровья [25]; и хотя испытания на безопасность были успешными, подходы с инфузией молодой крови не продемонстрировали свою эффективность в улучшении здоровья мозга или любой другой ткани в клинике [26]. Вместе с тем, эксперименты по гетерохронной гемотрансфузии показали, что в отсутствие обобществления органов и обогащения окружающей среды, как при парабиозе, молодая кровь не омолаживает старый мозг [21].
По мере того, как мы исследуем и формируем эволюционно консервативную парадигму системного омоложения, наши данные демонстрируют, что молодая кровь не является основным фактором, и вместо этого разбавление старой плазмы крови дает надежное переключение системной сигнальной среды на молодость и здоровье, омолаживая различные ткани. [18]. В этом отчете исследование мозга ограничивалось нейрогенезом гиппокампа; здесь мы расширяем работу на другие важные аспекты здоровья мозга: нейровоспаление и когнитивные способности. Наши данные демонстрируют, что нейровоспаление (в частности, активация микроглии) снижается, а когнитивные способности старых мышей (тесты на новый объект и на новую текстуру) улучшаются после одного NBE. В корреляции с фенотипами здоровья и функции омоложенного мозга наш сравнительный протеомный анализ выявил специфические нейропротекторные, нейрогенные и регулирующие нейроактивность белки, которые становятся системно активными у мышей и людей после разбавления плазмы. Учитывая, что терапевтический плазмообмен (TPE) одобрен FDA, это исследование предлагает использовать эту процедуру для предотвращения, ослабления и, возможно, даже обращения вспять дегенеративных и воспалительных заболеваний мозга.
Полученные результаты
Когнитивные способности старых мышей улучшаются с помощью одной процедуры NBE
Молодые (2–4 месяца) и старые (22–24 месяца) самцы мышей C57 / B6 прошли одну процедуру NBE, как опубликовано [18]. Изохронные обмены между молодыми мышами (молодые меняются с молодыми, YY) и старыми мышами (старые меняются со старыми, OO) были выполнены в качестве контроля процедуры. Через шесть дней после однократного NBE или контрольного обмена крови YY или OO были выполнены анализы нейровоспаления головного мозга, тесты на когнитивные способности и анализы протеомики крови, как в [21, 27, 28, 29] (рис. 1a, b).
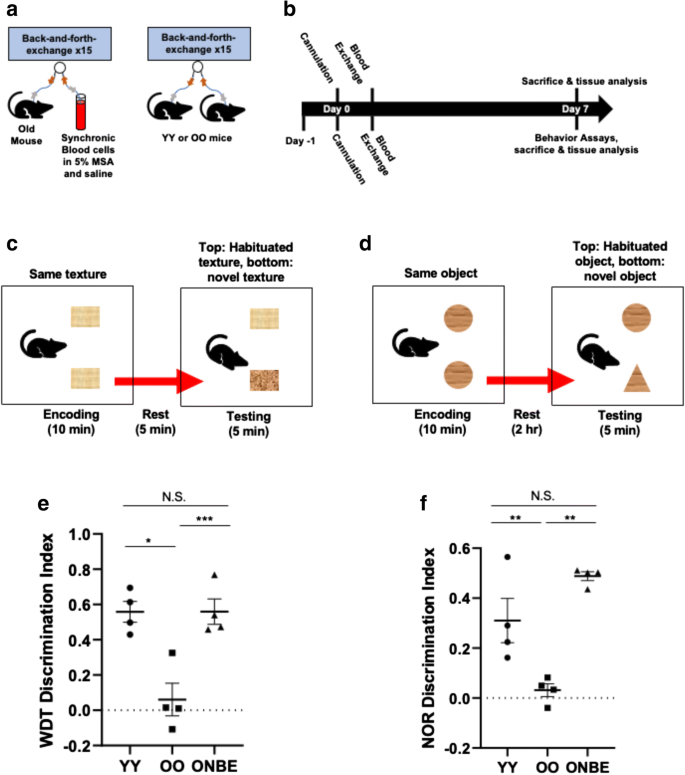 Рисунок 1. NBE быстро и надежно улучшает когнитивные способности старых мышей. Схема эксперимента. Катетеры были установлены в яремные вены молодых или старых мышей, у которых 50% плазмы крови заменено физиологическим раствором (0,9% хлорид натрия), 5% сывороточным альбумином мыши (MSA) и синхронными клетками крови, как было опубликовано ранее [ 21]. b Хронология. В день 1 мышей приучали к поведенческим тестам на распознавание усиками (WD) и распознавание новых объектов (NOR). Этим мышам была проведена канюляция яремной вены в день 0 и обмен крови в день 1. Анализы WD и NOR были выполнены в день 6. Образцы крови и головного мозга были собраны для анализа ткани. c Схемы мышей, выполняющих задачу распознавания усиками и d тесты распознавания новых объектов. e Мыши OO показали низкие результаты по сравнению с мышами YY, как и ожидалось (* значение p = 0,024). Мыши ONBE были намного лучше в различении между привычной и новой текстурой по сравнению с животными OO (*** значение p <0,00001). Интересно, что мыши ONBE могли различать новые и привычные текстуры так же эффективно, как и мыши YY (N.S. значение p = 0,01). f Аналогичные тенденции наблюдались в исследованиях распознавания новых объектов. YY по сравнению с ONBE N.S. Значение p = 0,99, YY по сравнению с OO ** значение p <0,004, OO по сравнению с ONBE ** значение p <0,006. Значения p были получены с помощью одностороннего критерия t Стьюдента, например, как типичный для оценки степени различий средних значений из трех независимых групп лечения, а не сравнения каждой группы с другой. N для YY = 4, N для OO = 4 и N для ONBE = 4 для каждого поведенческого анализа.
Рисунок 1. NBE быстро и надежно улучшает когнитивные способности старых мышей. Схема эксперимента. Катетеры были установлены в яремные вены молодых или старых мышей, у которых 50% плазмы крови заменено физиологическим раствором (0,9% хлорид натрия), 5% сывороточным альбумином мыши (MSA) и синхронными клетками крови, как было опубликовано ранее [ 21]. b Хронология. В день 1 мышей приучали к поведенческим тестам на распознавание усиками (WD) и распознавание новых объектов (NOR). Этим мышам была проведена канюляция яремной вены в день 0 и обмен крови в день 1. Анализы WD и NOR были выполнены в день 6. Образцы крови и головного мозга были собраны для анализа ткани. c Схемы мышей, выполняющих задачу распознавания усиками и d тесты распознавания новых объектов. e Мыши OO показали низкие результаты по сравнению с мышами YY, как и ожидалось (* значение p = 0,024). Мыши ONBE были намного лучше в различении между привычной и новой текстурой по сравнению с животными OO (*** значение p <0,00001). Интересно, что мыши ONBE могли различать новые и привычные текстуры так же эффективно, как и мыши YY (N.S. значение p = 0,01). f Аналогичные тенденции наблюдались в исследованиях распознавания новых объектов. YY по сравнению с ONBE N.S. Значение p = 0,99, YY по сравнению с OO ** значение p <0,004, OO по сравнению с ONBE ** значение p <0,006. Значения p были получены с помощью одностороннего критерия t Стьюдента, например, как типичный для оценки степени различий средних значений из трех независимых групп лечения, а не сравнения каждой группы с другой. N для YY = 4, N для OO = 4 и N для ONBE = 4 для каждого поведенческого анализа.
Когнитивные изменения происходят естественным образом в процессе старения млекопитающих и обычно связаны с ухудшением обучаемости и памяти [30]. Пожилые млекопитающие демонстрируют плохую когнитивную способность обнаруживать новые текстуры и новые объекты и взаимодействовать с ними, по сравнению с гораздо большей любознательностью в таких условиях у молодых млекопитающих [9, 10, 15, 29, 31, 32]. Этот аспект познания, который также отражает качество кратковременной памяти, может быть исследован в задаче распознавания усиками (WD), которая проверяет сенсорную обработку через бочкообразную кору и память через гиппокамп [32, 33], а также в тесте на распознавание новых объектов (NOR) [28], при котором сенсорная информация и информация из памяти обрабатываются как через гиппокамп, так и через периринальные области [34]. У пожилых мышей обычно наблюдается дефицит кратковременной памяти, что усугубляет их неспособность различать разные текстуры и разные объекты [29].
Тесты на распознавания усиками (рис. 1c) и распознавание новых объектов (рис. 1d) были выполнены после NBE на старых мышах, как в [29], с использованием мышей с изохронным обменом OO в качестве контроля процедуры и мышей с изохронным обменом YY в качестве стандарта здоровых молодых познавательных способностей. Как и ожидалось, у мышей OO наблюдалось сильное возрастное снижение показателей тестов NOR и WD по сравнению с животными YY (рис. 1e, f). Однако однократное разведение старой плазмы привело к значительному улучшению когнитивных функций старых мышей, которые фактически стали аналогичными когорте YY (рис. 1e и f). Фаза кодирования для каждого из этих поведенческих исследований показана на дополнительном рисунке 1.
Эти результаты показывают, что функциональность старого мозга (кратковременная память и любознательность) значительно и быстро улучшается за счет разбавления старой плазмы крови.
Возрастное увеличение нейровоспаления значительно ослабляется NBE
Нейровоспаление увеличивается с возрастом у мышей и людей, что способствует снижению когнитивных способностей у старых млекопитающих [9, 15, 29]. Таким образом, мы изучили, коррелируют ли когнитивные улучшения у старых мышей после NBE со снижением нейровоспаления.
С этой целью мы выполнили иммунофлуоресценцию на CD68, маркере активированной микроглии, в серийных криосрезах мозга молодых и старых мышей размером 25 мкм через 6 дней после изохронного обмена NBE или OO, а также в качестве положительного контроля для слабого нейровоспаления, обмен YY мышей.
На рис. 2а показана анатомическая карта, изображающая различные структуры в коронарных срезах мозга мыши (адаптированные из Атласа мозга Аллена), которые были исследованы на наличие клеток CD68 +. Срезы собирали с плоскости максимально обнаженной зубчатой извилины (ЗГ) гиппокампа. Активированные CD68 + (высокие) микроглиальные клетки были обнаружены непосредственно под (вентральной) по отношению к DG-части мозга, в таламусе, среднем мозге и внутренней зоне таламуса в головном мозге OO. Эти области разрезов были отображены; и другие части мозга OO также были профилированы, но не имели обнаруживаемых микроглиальных клеток CD68 +. Срезы мозга YY и ONBE изучались аналогично, например, по всему мозгу в последовательных 25-мкм криосрезах.
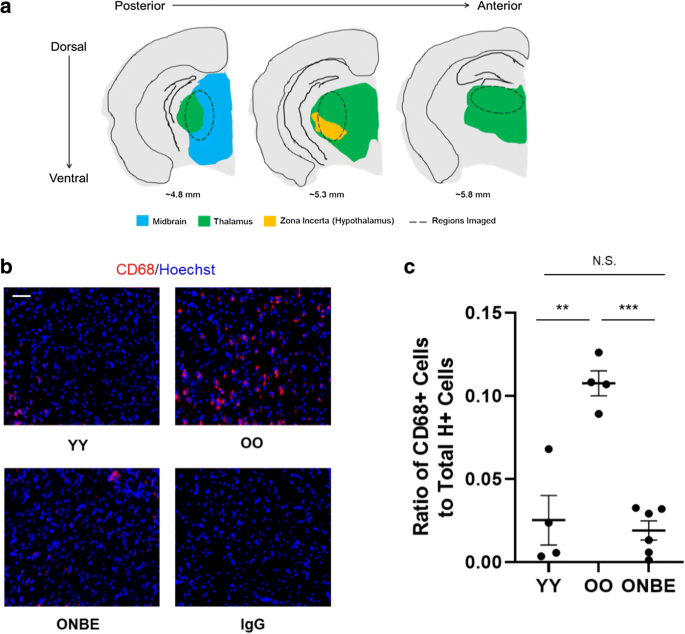 Рисунок 2. У старых мышей нейровоспаление снижается с помощью одного NBE. a Схема серийных срезов 25 мкм (обозначенных пунктирными овалами), которые были взяты из областей вентральных к зубчатой извилине, например, где было обнаружено очевидное возрастное увеличение относительного количества клеток CD68 + (зеленый: таламус, синий: средний мозг и оранжевый: внутренняя зона таламуса). b Иммунофлуоресценцию проводили для анализа CD68-положительной (красной) активированной микроглии в областях таламуса / гипоталамуса / среднего мозга мышей каждой когорты. Типичные двойные положительные клетки CD68 / Hoechst в указанных областях показаны для мышей YY, OO (изохронные контроли) и ONBE. Отрицательные контроли IgG, соответствующие изотипу, показывают отсутствие неспецифической флуоресценции. Масштабная линейка 50 мкм. c Количественная оценка относительной частоты активированной микроглии CD68 + / Hoechst + в таламусе. Нейровоспаление значительно снижено у мышей ONBE по сравнению с таковым у мышей OO (*** значение p <0,00002). Относительные количества активированной микроглии существенно не различаются между мышами YY и мышами ONBE (значение N.S. p = 0,27). ** значение p YY по сравнению с OO <0,003. Значения p были получены с помощью двустороннего критерия Стьюдента. N YY = 4, N OO = 4, N ONBE = 7
Рисунок 2. У старых мышей нейровоспаление снижается с помощью одного NBE. a Схема серийных срезов 25 мкм (обозначенных пунктирными овалами), которые были взяты из областей вентральных к зубчатой извилине, например, где было обнаружено очевидное возрастное увеличение относительного количества клеток CD68 + (зеленый: таламус, синий: средний мозг и оранжевый: внутренняя зона таламуса). b Иммунофлуоресценцию проводили для анализа CD68-положительной (красной) активированной микроглии в областях таламуса / гипоталамуса / среднего мозга мышей каждой когорты. Типичные двойные положительные клетки CD68 / Hoechst в указанных областях показаны для мышей YY, OO (изохронные контроли) и ONBE. Отрицательные контроли IgG, соответствующие изотипу, показывают отсутствие неспецифической флуоресценции. Масштабная линейка 50 мкм. c Количественная оценка относительной частоты активированной микроглии CD68 + / Hoechst + в таламусе. Нейровоспаление значительно снижено у мышей ONBE по сравнению с таковым у мышей OO (*** значение p <0,00002). Относительные количества активированной микроглии существенно не различаются между мышами YY и мышами ONBE (значение N.S. p = 0,27). ** значение p YY по сравнению с OO <0,003. Значения p были получены с помощью двустороннего критерия Стьюдента. N YY = 4, N OO = 4, N ONBE = 7
Как показано на рис. 2b и количественно представлено на с, CD68 + -клетки были многочисленны в старом контрольном мозге (OO) и минимальны / отсутствовали в молодом контрольном мозге (YY), что согласуется с ранее опубликованным возрастным увеличением CD68 + резидентных клеток [10, 17, 29]. Важно отметить, что нейровоспаление значительно уменьшилось у старых мышей после одного NBE. Репрезентативные и количественные изображения относятся к областям, показанным на рис. 2а; профилирование всего мозга YY и ONBE с помощью иммунофлуоресцентной микроскопии также не выявило клеток CD68 + в других местах. Неспецифическая иммунофлуоресценция совпадающих по изотипу контролей IgG была незначительной (рис. 2b).
Эти результаты демонстрируют, что нейровоспаление значительно и быстро уменьшается в мозге старых мышей после однократного разведения большого объема плазмы крови с помощью NBE.
ABT 263 сенолитик и NBE оба уменьшают старение мозга, но ABT 263 не может усилить нейрогенез или значительно уменьшить нейровоспаление у старых мышей
Чтобы изучить, могут ли и в какой степени положительные эффекты NBE эмулироваться за счет удаления стареющих клеток, мы провели исследования с ABT 263.
ABT 263 (Navitoclax) - химиотерапевтический препарат, который используется для индукции апоптоза в клетках рака шейки матки, пищевода, лейкемии и рака легких путем ингибирования антиапоптотических белков, Bcl-2 и Bcl-xL [35,36,37,38] . ABT 263, как было показано, избирательно очищает стареющие клетки in vivo посредством ингибирования белков Bcl [39,40,41] и снижает SASP [42].
Лекарства ABT были протестированы на глиобластомах пациентов в культуре [43]; но обычно считается, что эти молекулы слишком велики, чтобы пересечь гематоэнцефалический барьер (ГЭБ) [44]. Тем не менее, с возрастом ГЭБ становится более проницаемым даже для таких крупных белков, как альбумин [45,46,47]. Кроме того, сообщалось, что периферическое воспаление влияет на центральное воспаление и на здоровье мозга [48,49,50]. Белки периферического системного, связанного со старением секреторного фенотипа (SASP) проходят через стареющий ГЭБ и вызывают нейровоспаление и иммунную инфильтрацию [45, 46, 51, 52].
Поэтому мы предположили, что удаление периферических стареющих клеток с помощью ABT 263 может системно снижать уровни SASP, предотвращая их проникновение в мозг, тем самым улучшая нейрогенез гиппокампа, и ослабляя нейровоспаление у старых мышей. И нам было интересно определить, может ли разбавление системного SASP с помощью NBE уменьшить нагрузку на стареющие клетки в старом мозге.
Чтобы проверить эти гипотезы, мы сравнили эффекты ABT 263 и NBE, используя анализ SA-βGal на срезах мозга. ABT 263 или контрольный носитель вводили старым мышам C57 / B6 (возраст 22-24 месяца) через желудочный зонд в течение двух 7-дневных циклов с 2-недельным интервалом времени между циклами из расчета 50 мг на кг массы тела в день. NBE и контрольный изохронный OO-обмен выполняли, как описано выше. Свежезамороженные 25-мкм срезы головного мозга были проанализированы окрашиванием связанной со старением β-галактозидазой (SA-β-gal), при этом обнаружилось, что как ABT 263, так и NBE значительно снижали нагрузку стареющих клеток в старом мозге по сравнению с контролем для ABT 263 и контролем обмена OO для NBE (рис. 3a, b).
Fig. 3
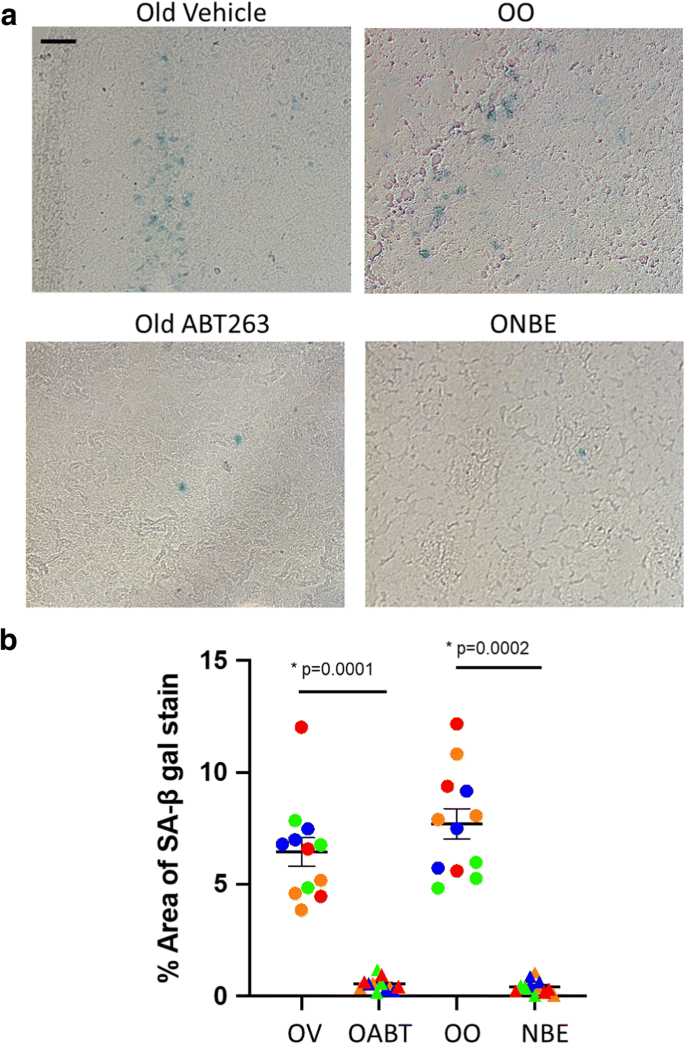 Рисунок 3. BT 263 и NBE снижают старение мозга. Свежезамороженные 25-мкм криосрезы старого мозга анализировали с помощью SA-βGal в соответствии с инструкциями производителя (9860S, Cell Signaling Technology). Репрезентативные изображения окрашивания SA-β-gal, масштабная линейка = 50 мкм. b Процент SA-β-gal-положительных областей на срез был количественно определен и сравнен между когортами. Старение старого мозга было уменьшено как ABT 263 (ABT), так и NBE по сравнению с контрольным носителем (OV) и изохронным обменом старой крови на старый (OO). Значения p были получены с помощью двустороннего критерия Стьюдента. ABT 263 по сравнению с транспортным средством p = 0,0001, NBE по сравнению с OO p = 0,0002. N = 4 животных каждой когорты. Цветовое кодирование показывает распределение SA-βGal-положительных клеток в серийных срезах мозга каждого исследованного животного, где один и тот же цвет представляет серийные срезы одного и того же мозга (круги, OV или OO; треугольники, ABT 263 или NBE)
Рисунок 3. BT 263 и NBE снижают старение мозга. Свежезамороженные 25-мкм криосрезы старого мозга анализировали с помощью SA-βGal в соответствии с инструкциями производителя (9860S, Cell Signaling Technology). Репрезентативные изображения окрашивания SA-β-gal, масштабная линейка = 50 мкм. b Процент SA-β-gal-положительных областей на срез был количественно определен и сравнен между когортами. Старение старого мозга было уменьшено как ABT 263 (ABT), так и NBE по сравнению с контрольным носителем (OV) и изохронным обменом старой крови на старый (OO). Значения p были получены с помощью двустороннего критерия Стьюдента. ABT 263 по сравнению с транспортным средством p = 0,0001, NBE по сравнению с OO p = 0,0002. N = 4 животных каждой когорты. Цветовое кодирование показывает распределение SA-βGal-положительных клеток в серийных срезах мозга каждого исследованного животного, где один и тот же цвет представляет серийные срезы одного и того же мозга (круги, OV или OO; треугольники, ABT 263 или NBE)
Таким образом, что интересно, сенолитики периферического действия и разжижение старой системной среды уменьшают старение мозга.
Предыдущие исследования показали, что повреждение мышц инъекциями кардиотоксина (CTX) ухудшает нейрогенез гиппокампа у мышей [21, 29]. Чтобы исследовать эти эффекты в контексте введения ABT 263, некоторые мыши получили повреждение CTX на 30 день лечения, в то время как другие нет (рис. 4a). Нейрогенез и нейровоспаление были исследованы в головном мозге этих животных, как опубликовано [18, 21, 29] и показано на рис. 2.
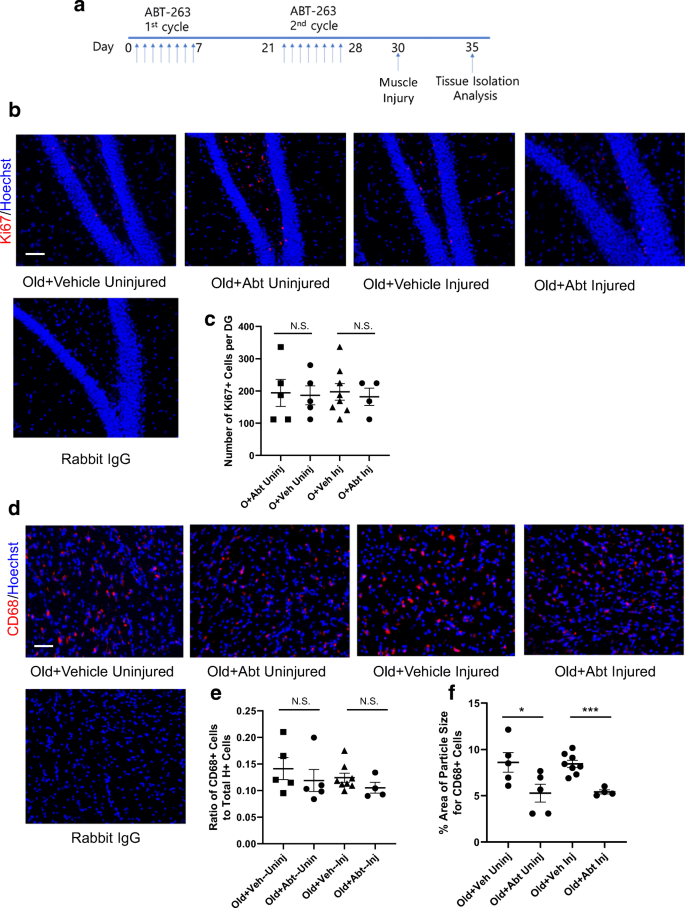 Рисунок 4. Эффекты сенолитика ABT 263 на нейрогенез гиппокампа и нейровоспаление старых мышей. Схема исследования. Было два 7-дневных периода, когда мышам вводили ABT 263 или носитель через желудочный зонд один раз в день. Между 7-дневными периодом желудочного зондирования следовал 2-недельный интервал. В ТА-мышцы некоторых мышей вводили кардиотоксин для экспериментального повреждения, в то время как другие животные не были повреждены. b Мозги были мгновенно заморожены и подвергнуты серийному криосрезу 25 мкм. Эти срезы подвергали иммуноанализу с использованием антител против Ki67 (маркер пролиферации) с использованием Hoechst для контрастного окрашивания всех ядер. Репрезентативные изображения зубчатой извилины гиппокампа показывают двойные положительные клетки Ki67 (красный) / Hoechst (синий) в субгранулярной зоне (пролиферирующие NPC SGZ). c Количественное определение Ki-67 + / Hoechst + SGZ клеток в зубчатой извилине было выполнено как для травмированных, так и для неповрежденных когорт. ABT 263 не улучшал нейрогенез гиппокампа ни в одной из групп. Old + Veh Uninj в сравнении с O + Abt Uninj значение p = 0,89; Old + Veh Inj по сравнению с O + Abt Inj значение p = 0,72. Значения p были получены с помощью двустороннего критерия Стьюдента. N Old + Veh Uninj = 5, N Old + Abt Uninj = 5, N Old + Veh Inj = 8, N Old + Abt Inj = 4. d Иммунофлуоресценцию проводили для CD68 (активированный маркер микроглии). Репрезентативные изображения двойных положительных клеток CD68 (красный) / Hoechst (синий). e Количественная оценка частоты клеток CD68 + / Hoechst + в головном мозге была выполнена для когорт, травмированных и здоровых. ABT 263 действительно снижал частоту клеток CD68 +. Old + Veh Uninj в сравнении с O + Abt Uninj значение p = 0,47; Old + Veh Inj по сравнению с O + Abt Inj значение p = 0,21. Н.С. незначительный. f Размер кластера CD68 на клетку анализ проводили на Фиджи. Размер кластеров CD68 был значительно уменьшен ABT 263. Old + Veh Uninj по сравнению с O + Abt Uninj значение p = 0,05; Old + Veh Inj по сравнению с O + Abt Inj значение p <0,0005. Значения p были получены с помощью двустороннего критерия Стьюдента. Для всех экспериментов: N старого + Veh Uninj = 5, N старого + Abt Uninj = 5, N старого + Veh Inj = 8, N старого + Abt Inj = 4. Шкала шкалы = 50 мкм. Не было неспецифической флуоресценции в отрицательных контролях IgG, соответствующих изотипу.
Рисунок 4. Эффекты сенолитика ABT 263 на нейрогенез гиппокампа и нейровоспаление старых мышей. Схема исследования. Было два 7-дневных периода, когда мышам вводили ABT 263 или носитель через желудочный зонд один раз в день. Между 7-дневными периодом желудочного зондирования следовал 2-недельный интервал. В ТА-мышцы некоторых мышей вводили кардиотоксин для экспериментального повреждения, в то время как другие животные не были повреждены. b Мозги были мгновенно заморожены и подвергнуты серийному криосрезу 25 мкм. Эти срезы подвергали иммуноанализу с использованием антител против Ki67 (маркер пролиферации) с использованием Hoechst для контрастного окрашивания всех ядер. Репрезентативные изображения зубчатой извилины гиппокампа показывают двойные положительные клетки Ki67 (красный) / Hoechst (синий) в субгранулярной зоне (пролиферирующие NPC SGZ). c Количественное определение Ki-67 + / Hoechst + SGZ клеток в зубчатой извилине было выполнено как для травмированных, так и для неповрежденных когорт. ABT 263 не улучшал нейрогенез гиппокампа ни в одной из групп. Old + Veh Uninj в сравнении с O + Abt Uninj значение p = 0,89; Old + Veh Inj по сравнению с O + Abt Inj значение p = 0,72. Значения p были получены с помощью двустороннего критерия Стьюдента. N Old + Veh Uninj = 5, N Old + Abt Uninj = 5, N Old + Veh Inj = 8, N Old + Abt Inj = 4. d Иммунофлуоресценцию проводили для CD68 (активированный маркер микроглии). Репрезентативные изображения двойных положительных клеток CD68 (красный) / Hoechst (синий). e Количественная оценка частоты клеток CD68 + / Hoechst + в головном мозге была выполнена для когорт, травмированных и здоровых. ABT 263 действительно снижал частоту клеток CD68 +. Old + Veh Uninj в сравнении с O + Abt Uninj значение p = 0,47; Old + Veh Inj по сравнению с O + Abt Inj значение p = 0,21. Н.С. незначительный. f Размер кластера CD68 на клетку анализ проводили на Фиджи. Размер кластеров CD68 был значительно уменьшен ABT 263. Old + Veh Uninj по сравнению с O + Abt Uninj значение p = 0,05; Old + Veh Inj по сравнению с O + Abt Inj значение p <0,0005. Значения p были получены с помощью двустороннего критерия Стьюдента. Для всех экспериментов: N старого + Veh Uninj = 5, N старого + Abt Uninj = 5, N старого + Veh Inj = 8, N старого + Abt Inj = 4. Шкала шкалы = 50 мкм. Не было неспецифической флуоресценции в отрицательных контролях IgG, соответствующих изотипу.
Нейрогенез происходит в субгранулярной зоне (SGZ) DG, снижается с возрастом и, как было показано, находится под влиянием системных факторов [5, 9, 10, 18, 21, 29]. Пролиферацию нейральных клеток-предшественников (NPC), которые рождаются в SGZ DG, анализировали с помощью иммунофлуоресценции маркера пролиферации Ki67 в серийных срезах мозга (фиг. 4b). Нейрогенез количественно оценивали по толщине каждого гиппокампа (рис. 4c). В соответствии с плохим нейрогенезом гиппокампа, который типичен для старых мышей, животные в контрольной группе имели ~ 150-200 NPC на DG [18, 21, 29]. Интересно отметить, что количество пролиферирующих NPC в гиппокампе старых мышей, которых лечили ABT 263, существенно не отличалось от такового у контрольных носителей. Количество пролиферирующих NPC в SGZ старых мышей, получавших ABT 263– и контрольных старых мышей было таким же, как и в когорте OO, и все они были значительно ниже, чем в группе ONBE (дополнительный рисунок 2A).
Степень нейровоспаления оценивали с использованием CD68 в качестве маркера активированной микроглии (как на рис. 2а). Относительная частота клеток CD68 + у старых мышей, получавших ABT 263, оставалась такой же высокой, как и в контрольной группе (рис. 4d, e). Частота клеток CD68 + была сходной между когортами ABT 263 и контрольным носителем и когортой OO, и все они были значительно выше по сравнению с частотой клеток CD68 + в когорте ONBE (дополнительный рисунок 2B).
При проведении этих исследований мы заметили, что величина флуоресценции CD68 оказалась ниже в когорте ABT 263, чем в контрольной группе (даже несмотря на то, что количество клеток CD68 + не изменилось). Количественная оценка размера кластеров CD68 на клетку подтвердила это наблюдение и продемонстрировала значительное уменьшение размера кластеров CD68 при лечении старых мышей ABT 263 (рис. 4f).
Эти результаты демонстрируют, что по сравнению с NBE сенолитический ABT 263 не усиливает нейрогенез в гиппокампе и имеет более слабый, но измеримый эффект на ослабление нейровоспаления. Эти различия проявляются, хотя и ABT 263, и NBE уменьшают старение мозга и системно ослабляют SASP.
Эволюционно консервативные эффекты NBE и TPE на системные белки с прямым влиянием на мозг
Учитывая выявленные множественные положительные эффекты NBE на здоровье и функции мозга и постулируемые омолаживающие эффекты TPE, мы провели сравнительный протеомный анализ сыворотки крови (мыши и человека), которые были изолированы до и после этих процедур (через 6 дней после NBE и через 1 месяц после TPE). Как упоминалось выше, с возрастом ГЭБ становится проницаемым для ряда системных белков (включая лиганды TGF-бета и альбумин [45,46,47]), что свидетельствует о повышенной способности периферических белков действовать в старом мозге.
Начав с протеомики системных факторов, которые модулировались NBE или TPE и показали> 2-кратное изменение при p <0,05, мы сосредоточились на сравнительных уровнях белков, которые непосредственно регулируют здоровье мозга, включая рецепторы, присутствующие на нейронах, микроглии, или других резидентных клетках мозга; лиганды, влияющие на пролиферацию и / или дифференцировку нервных клеток; и сигнальные белки, которые влияют на нейропластичность или являются нейропротекторными.
Эти исследования выявили 15 белков, которые имеют непосредственное значение для здоровья, поддержания и восстановления мозга и которые изменены в протеоме TPE, и 11 таких белков изменены в протеоме NBE мыши. Интересно, что у обоих видов разведение старой плазмы в конечном итоге привело к длительному повышению (а не снижению) этих детерминант, что согласуется с нашим общим выводом о восстановлении здорового состояния молекулярной сигнальной среды к (рис. 5).
Fig. 5
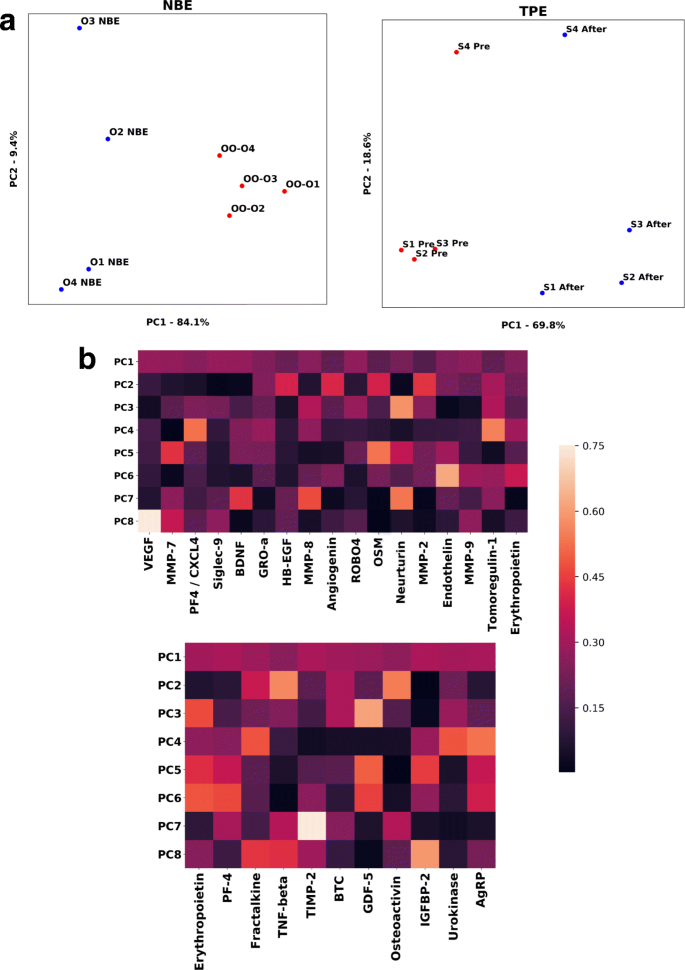 Рисунок 5. Множественные детерминанты здоровья и функции мозга одновременно повышаются после NBE и TPE. PCA был выполнен на совокупном наборе данных сывороточной протеомики контрольных старых мышей (старых заменяли на старые) и старых животных, которые прошли один цикл NBE (сыворотка была собрана через 6 дней после NBE), причем каждая точка представляла уникальную мышь, а значение вычислено преобразованием PCA с использованием одиннадцати белков, которые оказывают прямое влияние на здоровье мозга. В этой визуализации PCA агрегация протеома OO отделена от агрегации протеома ONBE. На первые два основных компонента приходится 93,5% различий между когортами. B. PCA проводили на протеомике сыворотки человека, сравнивая образцы Pre TPE с образцами, собранными через 1 месяц после одной процедуры, используя в качестве признаков 17 детерминант здоровья мозга. На первые два основных компонента приходится 88,4% вариации до и после лечения. C. Тепловые карты для оценок нагрузки для каждого основного компонента для 17 детерминант у человека (вверху) и 11 детерминант у мышей (внизу). Эритропоэтин и PF4 были эволюционно консервативными белками, на которые влияли NBE и TPE. Все образцы сыворотки были такими же, как в [18].
Рисунок 5. Множественные детерминанты здоровья и функции мозга одновременно повышаются после NBE и TPE. PCA был выполнен на совокупном наборе данных сывороточной протеомики контрольных старых мышей (старых заменяли на старые) и старых животных, которые прошли один цикл NBE (сыворотка была собрана через 6 дней после NBE), причем каждая точка представляла уникальную мышь, а значение вычислено преобразованием PCA с использованием одиннадцати белков, которые оказывают прямое влияние на здоровье мозга. В этой визуализации PCA агрегация протеома OO отделена от агрегации протеома ONBE. На первые два основных компонента приходится 93,5% различий между когортами. B. PCA проводили на протеомике сыворотки человека, сравнивая образцы Pre TPE с образцами, собранными через 1 месяц после одной процедуры, используя в качестве признаков 17 детерминант здоровья мозга. На первые два основных компонента приходится 88,4% вариации до и после лечения. C. Тепловые карты для оценок нагрузки для каждого основного компонента для 17 детерминант у человека (вверху) и 11 детерминант у мышей (внизу). Эритропоэтин и PF4 были эволюционно консервативными белками, на которые влияли NBE и TPE. Все образцы сыворотки были такими же, как в [18].
Интересно отметить, что белки, которые влияют на несколько параметров здоровья мозга, одновременно модулируются NBE и TPE, таким образом обеспечивая механизм для одновременного улучшения нескольких аспектов здоровья мозга: усиления функции мозга и синаптической пластичности, улучшения нейрозащиты, ослабления нейровоспаления, уменьшения нейротоксичности и усиления пролиферация и дифференцировка нейральных стволовых клеток (таблица 1).
|
|
Effects on the brain
|
|
TPE elevated
|
|
|
BDNF
|
Neurogenesis, neuroprotection, plasticity related to learning and memory [53]
|
|
Tomoregulin1
|
Neuroprotection: binds amyloid-β (Aβ) [54]
|
|
VEGF
|
Neuroprotection, secreted by neurons [55]
|
|
MMP-2, 7, 8, 9
|
Synaptic re-organization and memory [56]
|
|
Siglec
|
|
|
GRO-a
|
Oligodendrocyte proliferation and migration [59]
|
|
HB-EGF
|
Neuroprotection and function [60]
|
|
Angiogenin
|
Neuroprotection [61]
|
|
Robo4
|
Neurogenesis [62]
|
|
OSM
|
Neuroprotection and homeostasis of neural precursor cells [63]
|
|
Neurturin
|
Neuroprotection and neurogenesis [64]
|
|
Endothelin
|
Functional activity: neurotransmission [65]
|
|
NBE elevated
|
|
|
Platelet factor 4
|
Neurogenesis [66]
|
|
Erythropoietin
|
Neuroprotection [67]
|
|
Fractalkine
|
Homeostasis: neuron/glia crosstalk [68]
|
|
TNF-β
|
Neuronal plasticity, synaptic scaling, hippocampal neurogenesis [69]
|
|
TIMP-2
|
Inhibits neuronal proliferation and promotes the differentiation [70]
|
|
Betacellulin
|
|
|
GDF5
|
|
|
Osteoactivin
|
Neuroprotection: expressed on microglia—phagocytosis of myelin debris [75]
|
|
IGF-BP2
|
|
|
uPA
|
|
|
AgRP
|
|
Эти результаты подтверждают и экстраполируют положительные эффекты разведения старой плазмы на мозг, демонстрируя длительную рекалибровку системно присутствующих молекулярных факторов, имеющих прямое влияние на здоровье и функции мозга.
Обсуждение
Плазмообмен в настоящее время одобрен FDA для лечения других заболеваний, и, основываясь на нашей недавно опубликованной и текущей работе, TPE имеет большой терапевтический потенциал для лечения возрастных дегенеративных и воспалительных заболеваний мозга, а также общего возрастного ухудшения здоровья и функции мозга.
Использование TPE, при котором плазма пациента фильтруется и заменяется 5-20% раствором альбумина, было исследовано на предмет его эффективности для смягчения прогрессирования болезни Альцгеймера от легкой до умеренной. Исследование фазы I, начавшееся в 2005 г. и завершившееся в 2009 г., показало, что показатели познавательной способности и Aβ в спинномозговой жидкости изменились незначительно, однако объем гиппокампа, а также перфузия лобной и височной коры головного мозга увеличились за 6 месяцев наблюдения [82, 83].
Вторая фаза исследования, начавшаяся в 2007 г. и завершившаяся в 2017 г., включала более широкую популяцию пациентов, измеряли те же параметры, что и в фазе I, и при более интенсивном режиме плазмообмена. Результаты фазы II были замечательными в плане ослабления когнитивного снижения с доверительным интервалом, близким к 95%. Кроме того, у пациентов с плазмообменом наблюдалась стабилизация перфузии в лобных, височных и теменных областях [83,84,85]. Однако поведенческие, функциональные и когнитивные улучшения не были определены со статистической значимостью [86]. Текущие многоцентровые исследования проводятся для дальнейшего изучения эффективности TPE при лечении болезни Альцгеймера [83, 86].
Наши результаты согласуются с многообещающим направлением применения TPE для лечения болезни Альцгеймера; более того, наша работа расширяет омолаживающие фенотипы, предполагая, что заболевания мозга, как класс, и даже физиологическое старение мозга можно в какой-то момент предотвратить, ослабить и восстановить здоровье-молодость посредством соответствующего разбавления старой плазмы или старых факторов крови.
Кроме того, мы описываем системные белки-кандидаты, которые могут быть ответственны за положительные эффекты TPE / NBE на мозг, тем самым предлагая будущие терапевтические пути для использования агонистов / антагонистов в целях улучшения здоровья мозга и уменьшения старения мозга.
В этом отношении предыдущие исследования показали, что ослабление передачи сигналов TGF-бета ингибитором Alk5, в частности, в сочетании с эктопическим окситоцином, улучшает нейрогенез гиппокампа, уменьшает нейровоспаление, и улучшает познавательные способности у старых мышей [13, 29]. Эффекты NBE в целом сильнее, чем эффекты ингибитора Alk5 плюс окситоцин, что вполне ожидаемо, поскольку разведение старой плазмы действует по множеству механизмов и повышает уровни многочисленных «молодых» системных факторов [18].
Сравнение NBE с другим известным методом фармакологического омоложения, сенолитиками (ABT 263 (Navitoclax), которые вызывают апоптоз в стареющих клетках [87]), показало, что разбавление плазмы оказывает гораздо более сильное положительное влияние на старый мозг, чем удаление стареющих клеток.
Существует значительный интерес к разработке сенолитических методов лечения заболеваний, связанных с возрастом. ABT 263 представляет большой клинический интерес, поскольку изначально он использовался для лечения различных форм рака [88,89,90,91] и оказывает влияние на большинство периферических тканей, но для воздействия на опухоли головного мозга необходимо вводить дополнительные препараты [92, 93].
Тот факт, что NBE имеет более сильный эффект, чем ABT 263 при омоложении мозга, предполагает, что восстановление сигнальной среды для здоровья и молодости путем разбавления старой плазмы более надежно для омоложения мозга, чем ослабление SASP посредством удаления стареющих клеток. Интересно, что наши данные демонстрируют, что старение мозга снижается как периферически действующими ABT 263, так и NBE, в поддержку представления о том, что возрастное системное старение и SASP могут вызывать старение в ЦНС [45, 46, 51, 52]. Уменьшение размера кластера CD68, наблюдаемое в этой работе, может быть связано с умеренным ослаблением нейровоспаления [94] с помощью ABT 263. И один из предложенных механизмов улучшения болезни Альцгеймера действительно включает уменьшение нейровоспаления, когда периферическая модуляция влияет на воспаление центрального мозга [83]. Было бы интересно изучить, может ли ABT 263 быть эффективным в таких приложениях.
Недавние исследования показывают, что экспериментальные травмы головного мозга у мышей отрицательно влияют на здоровье гиппокампа (потеря синапсов, гибель нейронов) и влияют на познавательную способность, даже когда очаги таких повреждений и последующей активации микроглии находятся в другом месте мозга [95,96,97,98]. Более того, локальная активация микроглии приводит к широким воспалительным ответам в головном мозге за счет аутокринных и паракринных функций провоспалительного секретома, что кумулятивно увеличивает нейровоспаление во всем головном мозге [99, 100]. В соответствии с нашими выводами, травма таламуса (например, в одной из областей, где мы обнаружили что NEB уменьшает возрастное нейровоспаление) активирует микроглию и увековечивает нейродегенерацию и снижение когнитивных функций [101]. В совокупности эти результаты предполагают, что активация микроглии является диффузной и распространяется от одной области мозга к другой.
Наше исследование сопоставило возрастную частоту появления CD68 + клеток в таламусе, гипоталамусе и среднем мозге. Связанная с возрастом активация микроглии в каждой из этих областей действительно является причиной патологий головного мозга и, как ожидается, повлияет на функцию мозга и когнитивные способности [102,103,104,105]. И в целом нейровоспаление, по крайней мере частично, отвечает за возрастную потерю нейронов [106].
Следовательно, быстрое улучшение когнитивных функций у старых мышей в этом исследовании, как полагают, связано с устранением (посредством NBE) возрастной степени нейровоспаления. Следует отметить, что разведение плазмы также может иметь клиническое значение для ослабления гиперактивированной микроглии CD68 + при психических заболеваниях, когда нейровоспаление вовлечено в патологию [107].
Весьма маловероятно, что усиление нейрогенеза у старых мышей с помощью NBE [18] вносит вклад в наблюдаемое здесь улучшение когнитивных функций. Как и ранее [29], мы оценивали когнитивные способности через 1 неделю после однократной процедуры NBE; таким образом, не было достаточно времени для формирования новых нейронов и / или интеграции в сети.
В целом, эта работа поддерживает парадигму, согласно которой разбавление и восстановление системной сигнальной среды для здоровья и молодости способствует «молодым» факторам здоровья тканей, омолаживая мозг (и, фактически, все ткани и параметры, которые были изучены до настоящего времени [18 ]).
Материалы и методы
Животные
Все эксперименты и процедуры in vivo проводились в соответствии с политикой, установленной Управлением по уходу за лабораторными животными, и в соответствии с утвержденными протоколами Калифорнийского университета в Беркли и Института исследований старения Бака. Молодые самцы мышей C57BL / 6 (возраст 2 месяца) были приобретены в лаборатории Джексона, в то время как старые мыши (возраст 18 месяцев) были приобретены в Национальном институте старения (NIA). Старым мышам позволяли акклиматизироваться в том же помещении, где содержались молодые мыши, в течение нескольких недель перед исследованиями. Все мыши получали одинаковый рацион. Все старые мыши, использованные в когортах OO, ONBE, Old + Vehicle и Old + ABT 263, были получены из NIA.
Количество животных (N)
Анализ мощности был проведен для определения размеров выборки для представленных экспериментов, как описано ранее [18].
Операция по канюлированию яремной вены и процедуры обмена крови
Обратите внимание, что все оборудование, используемое для этих процедур, было стерилизовано в автоклаве. При повторном контакте с несколькими мышами использовали стерилизатор для шариков. Канюляцию яремной вены и процедуры афереза крови выполняли, как было опубликовано ранее [18, 21]. Вкратце, мышам давали бупренорфин (0,1 мг / кг) и анестезировали 1-3% изофлураном в кислороде до полного расслабления. На каждый глаз наносили офтальмологическую мазь для предотвращения высыхания. Мышей брили вокруг шеи, отдыхали в положении лежа на спине и три раза наносили хирургический скраб с бетадином на их голую кожу. Салфетки изопропанола использовали для удаления бетадина после каждого применения бетадина. Затем мышей помещали на стерильное поле. Когда не было реакции на защемление пальца, справа от средней линии делали разрез на 1–1,5 см и выделяли правую внутреннюю яремную вену. Шелковый шов 6-0 использовался для лигирования краниального конца вены. К лигированному концу вены прикладывали легкое натяжение, в то время как еще один шов 6-0 был проведен под веной и завязан свободным узлом. Для выполнения венотомии использовалась игла 25-го калибра со скошенным концом, загнутым наружу на 90 °. Конец 1-Fr предварительно гепаринизированного катетера 1–3-Fr (Instech Labs, C10PU-MJV1403) быстро вставляли в яремную вену, а каудальную лигатуру затягивали, чтобы удерживать катетер на месте. После подтверждения проходимости катетер закупоривали и накладывали дополнительную черепную лигатуру для фиксации катетера на месте. Затем мышей помещали в положение лежа на левом боку, чтобы продеть катетер между лопатками. Для этого использовались тупые щипцы, чтобы создать пространство под кожей, продвигая место разреза к лопаткам. Игла 16 калибра была помещена между лопатками и введена под кожу на уровне места разреза. Конец катетера 3-Fr пропущен через иглу калибра 16. Зажимы Reflex 7 использовались для закрытия места разреза, а катетер, выступающий за кожу, фиксировался каплей Dermabond. Мышей снимали с наркоза, вводили мелоксикам подкожно (5 мг / кг) в течение 7 дней после процедуры и давали им возможность восстановиться в клетке. На закрытый участок нанесена мазь с антибиотиком с лидокаином.
Кровь от молодых или старых мышей-доноров получали путем терминальной пункции сердца и антикоагулировали 3 единицами гепарина. Образцы крови центрифугировали при 500 g в течение 5 мин. Фракции плазмы, обогащенной тромбоцитами, были тщательно удалены; Осадки клеток крови ресуспендировали в физиологическом растворе и затем снова центрифугировали при 500 g в течение дополнительных 5 мин. Слой физиологического раствора удаляли, а затем осадки клеток крови ресуспендировали в равном объеме 5% MSA в нормальном физиологическом растворе и 0,9% хлористом натрия. По данным производителя, замещающий альбумин имеет чистоту> 95% (undefined). Эти смеси крови пропускали через пробирку с сетчатым колпачком FACS 50 мкм, чтобы отделить клетки и отфильтровать любые сгустки. Экстракорпоральный обмен крови выполняли между парами молодых мышей, парами старых мышей и молодых мышей или старых мышей и пробиркой, содержащей синхронные клетки крови в 5% MSA, через несколько часов после операции. Эти дизайнерские растворы крови были приготовлены непосредственно перед проведением обменов крови.
Обмен крови был выполнен через 24 часа после операции по канюлированию, чтобы обеспечить адекватное восстановление. Мышей еще раз анестезировали 1–3% изофлураном в кислороде и вводили офтальмологическую мазь в состоянии покоя в вентральном положении лежа. Катетеры оценивали на проходимость и промывали 3 единицами физиологического раствора гепарина. В общей сложности 150 мкл крови было заменено между мышью и пробиркой с нашим разработанным раствором крови 15 раз для 50% замены плазмы крови физиологическим раствором + альбуминовой жидкостью или для 50% синхронных обменов молодой или старой крови, как в [ 21]. Процесс обмена длится примерно 30–40 мин. После завершения катетеры были закупорены, мышей сняли с анестезии и дали им возможность восстановиться.
ABT-263 лечение
Для медикаментозного лечения самцов мышей C57BL / 6 в возрасте от 22 до 24 месяцев из Национального института старения (Bethesda, MD, США) лечили безлекарственным раствором или ABT 263 (APExBIO, США), разведенным в 10% этаноле, 30%. полиэтиленгликоль 400 и 60% Phosal 50 PG (Lipoid, Германия). ABT 263 вводили через желудочный зонд в дозе 50 мг на кг массы тела в день (мг / кг / день) в течение 7 дней на цикл в течение двух циклов с двухнедельным интервалом между циклами.
Кардиотоксиновая травма мышц
Мыши были травмированы внутримышечными инъекциями CTX (Sigma, 10 мл на мышцу при 0,1 мг / мл) в переднюю большеберцовую мышцу (TA) и икроножную мышцу (GA), как было опубликовано ранее [1, 18, 21, 29].
Тканевая изоляция
Мышей умерщвляли в соответствии с инструкциями администрации OLAC Калифорнийского университета в Беркли и Института Бака. Кровь собирали терминальной сердечной пункцией и оставляли для полного свертывания при комнатной температуре в течение не менее 30 минут. Образцы свернувшейся крови центрифугировали при скорости 5000 g в течение 5 мин для получения сыворотки. Произведено патологоанатомическое исследование головного мозга. Ткани помещали в Tissue-Tek с оптимальной температурой резки (OCT, Sakura Finetek, Нидерланды) и мгновенно замораживали в изопентане, охлажденном до -70 ° C с помощью сухого льда.
Разделение тканей и картирование мозга
Мозг с внедренной ОКТ был рассечен с помощью криостата. Корональные срезы толщиной 25 мкм прикрепляли к положительно заряженным предметным стеклам с добавлением золота. Криостат использовался для определения местоположения областей среднего мозга, таламуса и гипоталамуса, как указано выше. Эти области головного мозга располагались примерно в 4,5–4,8 мм от самого заднего края мозжечка. Последующие срезы ткани были взяты с пересечением на 1 мм этой отметки.
Окрашивание β-галактозидазой, ассоциированное со старением
Ассоциированное со старением окрашивание β-галактозидазой (SA-β-gal) выполняли с помощью коммерческого набора для окрашивания Senescence β-галактозидазой (9860S, Cell Signaling Technology) в соответствии с протоколом производителя. Вкратце, замороженные старые срезы мозга мышей фиксировали в фиксирующем растворе (входит в комплект) в течение 15 мин при комнатной температуре. После двукратной промывки PBS срезы окрашивали окрашивающим раствором β-галактозидазы при 37 ° C в течение ночи в сухом инкубаторе (Thermolyne, инкубатор типа I42300, США). После окрашивания изображения получали с помощью микроскопа Axio Imager A1 (Zeiss, Германия) при увеличении × 200. Процент стареющей области определяли путем деления окрашенных в синий цвет SA-β-gal-позитивных областей на общую площадь каждого раздела с помощью программного обеспечения ImageJ.
Антитела и реагенты для маркировки
Следующие антитела использовали в концентрации 0,5–1 мкг / мл:
CD68: Abcam, Rabbit, ab125212, 1:500
Ki67: Abcam, Rabbit, ab16667, 1:200
Isotype-matched IgGs: Sigma-Aldrich, Rabbit, 1:1000
Donkey anti-rabbit Alexa 546: Life Technologies, Invitrogen, Eugene, Oregon, A10040, lot #1946340, 1:2000
Для окрашивания ДНК использовали краситель Hoechst: Hoechst 33342, Sigma-Aldrich (B2261), 1: 1000.
Иммунофлуоресценция образцов мозга
Мозг мыши был серийно разрезан на срезы толщиной 25 мкм, которые затем фиксировали в 4% параформальдегиде в течение 4 мин при комнатной температуре. Затем эти срезы промывают 1 × фосфатно-солевым буфером (PBS) несколько раз (2–3 мин на одно полоскание) и затем проницают 0,1% тритоном-X на льду в течение 5 мин. Затем образцы промывали и блокировали PBS и 1% окрашивающим буфером 3 раза (2–3 мин на промывание). Затем образцы мозга обрабатывали первичными антителами и оставляли инкубироваться в течение ночи при 4 ° C. В течение следующего дня срезы промывают 3 раза (2–3 мин на одно полоскание) окрашивающим буфером, а затем покрывают вторичными антителами, как описано, и инкубируют в течение 2 часов. Затем образцы промывали 3 раза буфером для окрашивания (2–3 мин на полоскание); затем добавляли 2 капли монтажной среды Fluoromount (Sigma F4680) и поверх образцов помещали покровные стекла. Все срезы тканей для иммунофлуоресцентных исследований помещали на положительно заряженные золотые предметные стекла SuperFrost.
Поведенческие анализы
Тест на различение текстур усиками проводился, как описано ранее [27, 28], с фазами кодирования, покоя и тестирования, которые длились 10, 5 и 5 минут соответственно. Тест на распознавание нового объекта (NOR) также проводился, как описано ранее [27,28,29], с фазами кодирования, отдыха и тестирования, установленными на 10 минут, 2 часа и 10 минут соответственно. Такие модификации кодирования и продолжительности тестирования предназначены для учета медленного передвижения старых мышей. Поведенческие анализы проводились с аналитиком, который не имел информации о конкретных мышах (возраст, тип лечения).
Анализ основных компонентов
Анализ основных компонентов был выполнен посредством разложения по сингулярным значениям с помощью пакета декомпозиции Python-scikit-learn. Данные были масштабированы с помощью StandardScaler в методе предварительной обработки sklearn, а характеристики были нормализованы по средней экспрессии белка. Python-Matplotlib использовался для визуализации данных, преобразованных в PCA, а Python-Seaborn использовался для создания тепловых карт, используемых для визуализации загрузки PC для каждой функции белка.
Количественная оценка данных и статистика
Нейрогенез количественно оценивали путем подсчета количества клеток Ki67 + / H + в 200 мкм SGZ от каждой мыши из нескольких, как описано ранее [18, 29]. Нейровоспаление оценивали путем подсчета количества клеток CD68 + по отношению к общему количеству ядер, подсчитанному на поле зрения. Стратегии картирования были описаны на рис. 2. Все анализы проводились на срезах тканей, которые были отображены при увеличении × 20. Непарные, односторонние и двусторонние (в зависимости от ситуации) t-тесты Стьюдента были выполнены в GraphPad Prism 8 для всех данных анализа тканей.
Доступность данных
Все соответствующие данные были предоставлены при подаче этой рукописи.
Опубликовано: 15 ноября 2020 г.
Авторы: Мелод Мехдипур , Таха Мехдипур , Колин М. Скиннер , Натан Вонг , Чао Лю , Чиа-Чиен Чен , Ок, Хи Чон , И Цзо , Майкл Дж. Конбой и Ирина М. Конбой
Перевод Ник Сестрин
НОВОСТИ ПО ТЕМЕ:
Разведение плазмы улучшает когнитивные функции и снижает нейровоспаление у старых мышей
Наше недавнее исследование установило, что факторы молодой крови не являются причиной и не являются необходимостью для системного омоложения тканей млекопитающих...
Пора кончать со старой кровью - Джош Миттельдорф
2020 год обещает нам, что мы сможем сделать наши тела молодыми без явного восстановления молекулярных повреждений, но лишь просто изменив...
Омоложение тканей трех зародышевых листков путем замены плазмы старой крови солевым раствором альбумина
Аннотация Гетерохронный обмен крови омолаживает старые ткани, и большинство исследований о том, как это работает, фокусируется на молодой плазме, ее фракциях...
Обращение возраста: измерение эпигенетического возраста двух разных видов с помощью одних часов

Аннотация Известно, что молодая плазма крови оказывает благотворное влияние на различные органы у мышей. Однако не было известно, омолаживает ли молодая...
Прорыв в омоложении

Если вы избегаете громких заявлений и в течении длительного времени соблюдаете дисциплину недосказывания посреди яркого неонового мира, то возможно вы...
Эпидемия молодости: как прожить 120 лет и стать счастливым

Около 5% нынешних молодых и богатых проживут 120 лет и дольше, считают биохакеры. Читайте, что для этого нужно делать. Осенью 2017...
Близок ли конец старения - Интервью с Джорджем Чёрчем

Предисловие Грегори Фэйи Будучи биогеронтологом, я посещаю научные встречи, посвящённые старению, с начала 1980-х годов, и видел и слышал много потрясающих...
Почему Питер Тиль так интересуется молодой кровью

Более всего Питера Тиля, миллиардера-инвестора и основателя PayPal, интересует поиск способа избежать смерти. Он вкладывает миллионы в стартапы, работающие над...
Лекарство от старости скоро будет создано, но сможете ли вы его себе позволить?

Чувствуете? Это ваше тело медленно разрушается. Все мы стареем, и старение приводит болезни, которые в конечном счете большинство из нас...
Медик: "молодая" кровь снизила шансы развития рака и болезни Альцгеймера

Переливание крови молодых людей пожилым пациентам снизило вероятность развития рака и болезни Альцгеймера и привело к общему омоложению некоторых клеток и органов, сообщает сайт...
Биологи научились создавать кровь из стволовых клеток человека

Биологи из Бостона разработали методику превращения "перепрограммированных" стволовых клеток в заготовки клеток крови, что открывает дорогу для лечения болезней и создания "бесконечных" донорских запасов...
Пуповинная кровь человека омолодила мозг мышей, заявляют ученые

Переливание человеческой пуповинной крови в кровеносную систему пожилой мыши омолодило ее мозг и улучшило память, заявляют ученые в статье, опубликованной в журнале Nature. "Нейрофизиологи постоянно...
Выращивание клеток крови поставили на поток
Биологи придумали способ получать жизнеспособные эритроциты in vitro в количествах, пригодных для клинического применения. Для этого они создали «бессмертную» линию клеток-предшественников эритроцитов...
Центрифугу для типирования крови уместили во флешку
Китайские ученые разработали одноразовый экспресс-тест на бумажной основе, позволяющий определить группу крови человека с точностью более 99,9 процента. Результаты исследования...
Искусственная кровь
Биофизик Михаил Пантелеев о проблемах донорской крови, выращивании и программировании новых клеток из стволовых и прототипах искусственных тромбоцитов В настоящее время...
Клетки крови сделали из клеток кожи
Ученые из Сингапура научились делать клетки крови из клеток кожи. Они провели эксперимент на мышах – созданные таким образом клетки...
"Молодая" кровь не является эликсиром молодости, доказали ученые

Биологи из Калифорнии провели повторные опыты по переливанию крови молодых мышей в тело старых грызунов и не подтвердили того, что подобная процедура ведет к сколь-либо...
В США приготовились к испытаниям по омоложению людей плазмой крови
Специалисты из фирмы Ambrosia в Монтерее, штат Калифорния, готовятся к проведению первых испытаний по омоложению людей с помощью плазмы крови...
Советский кровезаменитель вновь начнут производить в России
«Голубая кровь», которую планируется использовать при пересадке головы, вновь появится на российском рынке. Кровезаменитель Перфторан, который планируют использовать во время первой...
Исцеление молодой кровью: продолжение
Гарвардские физиологи отбили атаку на «белок молодости» Ранее по теме: Учёные доказали, что обычное переливание крови может развернуть процесс старения вспять После...
О донорстве крови и костного мозга
«Донорство должно быть не только безвозмездным, но и добровольным» Фото: Oleg Pchelov / Shutterstock.com 17 июля 2015 года Читать еще: донорство крови, стволовые...
Великобритания начнёт клинические испытания искусственной крови уже в 2017 году
25 Июня 2015 Кровь для переливания – очень ценный продукт, постоянно необходимый медикам в больших количествах. Многочисленные пункты приёма крови зачастую...
Плазма крови
Пла́зма кро́ви (от греч. πλάσμα — нечто сформированное, образованное) — жидкая часть крови, в которой взвешены форменные элементы — вторая часть крови. Процентное...
Учёные доказали, что обычное переливание крови может развернуть процесс старения вспять
Учёные доказали, что обычное переливание крови может развернуть процесс старения вспять 6 Мая 2014 Две независимые группы учёных опубликовали результаты своих научных...