1.10.2014

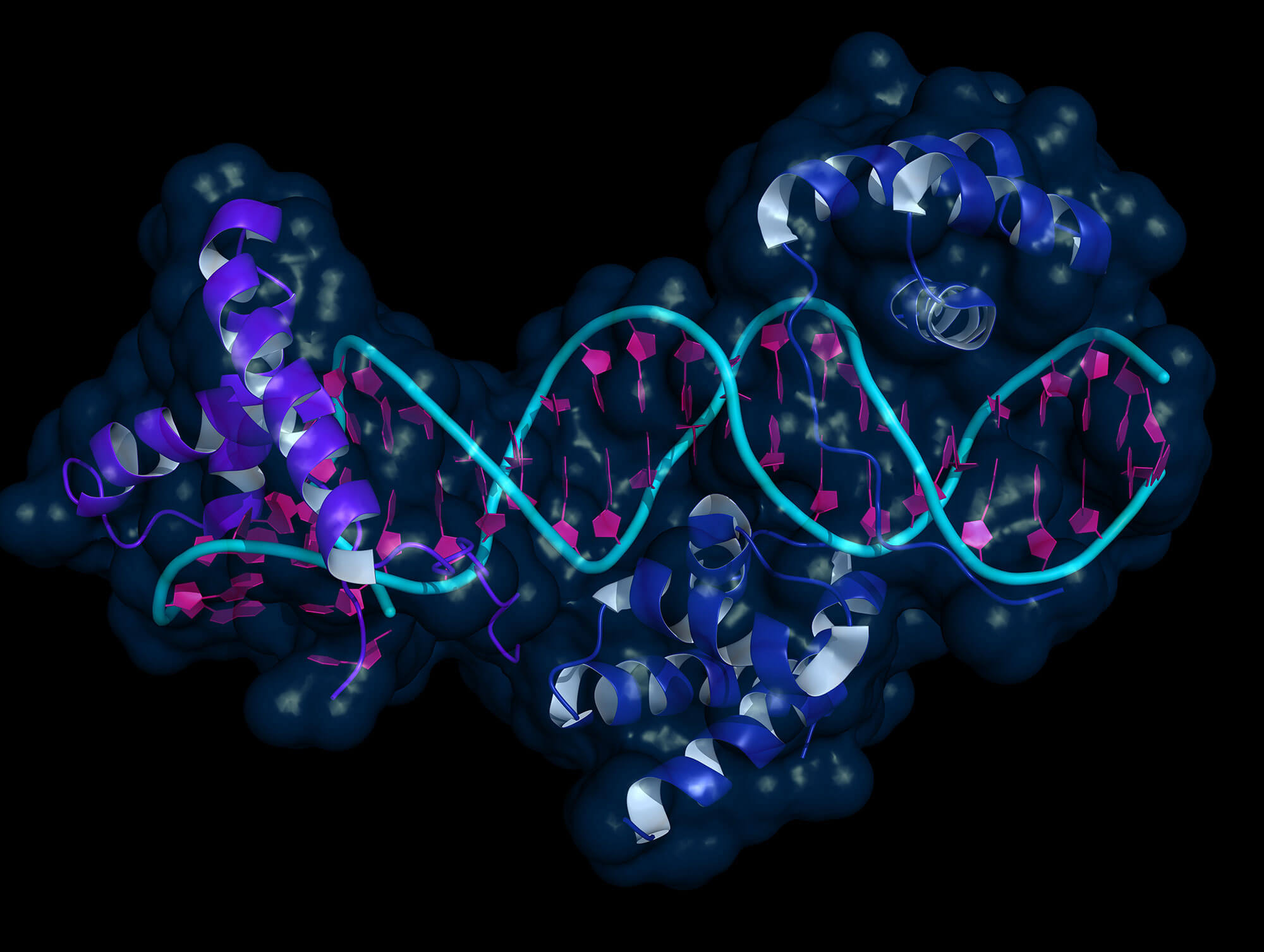
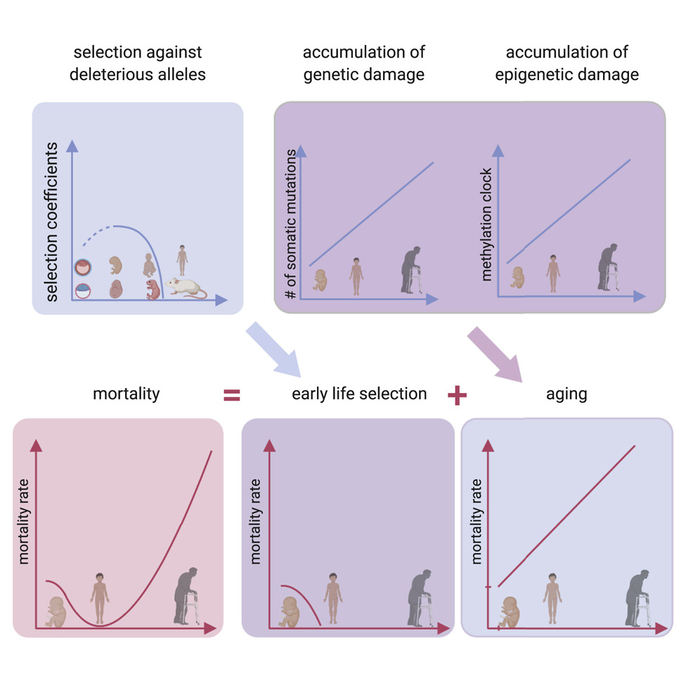
Рис. 1. Двухфазная регуляция дорзо-вентральной разметки нервной трубки (будущего спинного мозга). Показан поперечный срез нервной трубки на трех стадиях развития. На раннем этапе (1st phase. Morphogen-dependent) градиенты сигнальных белков — морфогенов BMP и SHH (показаны слева) — контролируют разделение прогениторных клеток (предшественников нейронов) на несколько слоев — доменов, из которых затем будут образовываться разные типы нейронов. Розовым цветом обозначена область предшественников мотонейронов (motorneurons (precursors)), голубым — дорзальных вставочных нейронов (dorsal interneurons). В дальнейшем начинается дифференцировка — превращение прогениторных клеток в нейроны, которые перестают делиться и мигрируют на периферию (показаныкосой штриховкой), а прогениторные нейроны остаются около невроцеля (вертикальная щель в центре каждого среза). На этом этапе градиенты BMP и SHH уже не влияют на пропорции и положение границ доменов, которые теперь регулируются в автономном режиме и определяются характерной для каждого домена скоростью превращения прогениторных клеток в нейроны. Схема из синопсиса к обсуждаемой статье.
Как в ходе развития эмбриона регулируются пропорции его быстро растущих частей — один из актуальных вопросов биологии развития. На примере дорзо-вентральной (спинно-брюшной) организации нервной трубки у мыши и курицы международный коллектив исследователей показал, что способ регуляции может меняться по мере роста органа. Первичное подразделение нервной трубки на участки с разными типами прогениторных клеток, из которых затем образуются разные типы нейронов, определяется градиентами сигнальных белков — морфогенов Shh, BMP и Wnt. Однако в дальнейшем, когда прогениторные клетки начинают дифференцироваться (превращаться в нейроны), влияние морфогенов резко снижается. На первый план выходит автономная регуляция, основанная на характерной для каждого типа прогениторных клеток скорости дифференцировки. Соотношение этих скоростей и определяемая ими дорзо-вентральная организация спинного мозга похожи у мыши и курицы, то есть эволюционно консервативны. Исследование проливает свет на то, каким образом могут сохраняться оптимальные пропорции частей организма при изменениях его размера в ходе эволюции.
Градиенты концентраций сигнальных белков — морфогенов, таких как Wnt, Shh или BMP, играют важную роль в «разметке» развивающегося эмбриона (и отдельных его частей) на дискретные области, клетки которых затем будут вести себя по-разному, формируя разные ткани, органы и отделы тела (см.: Как клетки понимают, что одни должны стать волосами, другие костями, третьи мозгами и т. п.?). Морфогены часто производятся особыми группами клеток (их иногда называют «организаторами», см. Эмбриональная индукция), откуда они распространяются путем диффузии или инициируя собственное производство в соседних клетках, причем концентрация морфогена падает по мере удаления от организатора.
От концентрации морфогенов в межклеточном пространстве зависит, какие наборы регуляторов следующего уровня — транскрипционных факторов — активируются внутри клеток. Транскрипционные факторы, в свою очередь, регулируют экспрессию множества генов, работа которых в конечном счете определяет судьбу клетки.
Градиенты морфогенов, определяющие активацию транскрипционных факторов, — не единственный механизм разметки (patterning) развивающегося эмбриона (см.: Идеи Алана Тьюринга помогли понять механизм развития пальцев у позвоночных, «Элементы», 18.12.2012). Многое в этих механизмах остается неясным. Например, не очень понятно, как регулируются пропорции быстрорастущих частей эмбриона. Ведь рост неизбежно ведет к изменению концентраций морфогенов в развивающихся тканях. Чтобы границы частей при этом не «поплыли», нужно либо динамически менять уровень производства морфогенов, подлаживаясь к меняющимся размерам подконтрольных зачатков, либо использовать другие способы регуляции. Эта проблема относится не только к эмбриональному росту, но и к изменению размеров тела в ходе эволюции. В некоторых случаях, как уже удалось выяснить, действительно происходит подстройка градиентов морфогенов (см.: T. Gregor et al., 2005. Diffusion and scaling during early embryonic pattern formation). Но всегда ли так бывает?
Исследование британских и австрийских биологов, результаты которого опубликованы в последнем выпуске журнала Science, дает отрицательный ответ на этот вопрос.
Авторы исследовали регуляцию дорзо-вентральной разметки нервной трубки мышиного эмбриона. Ранние стадии процесса изучены уже достаточно подробно. Крайне упрощенная схема показана на рис. 1, чуть более подробная — на рис. 2.

|
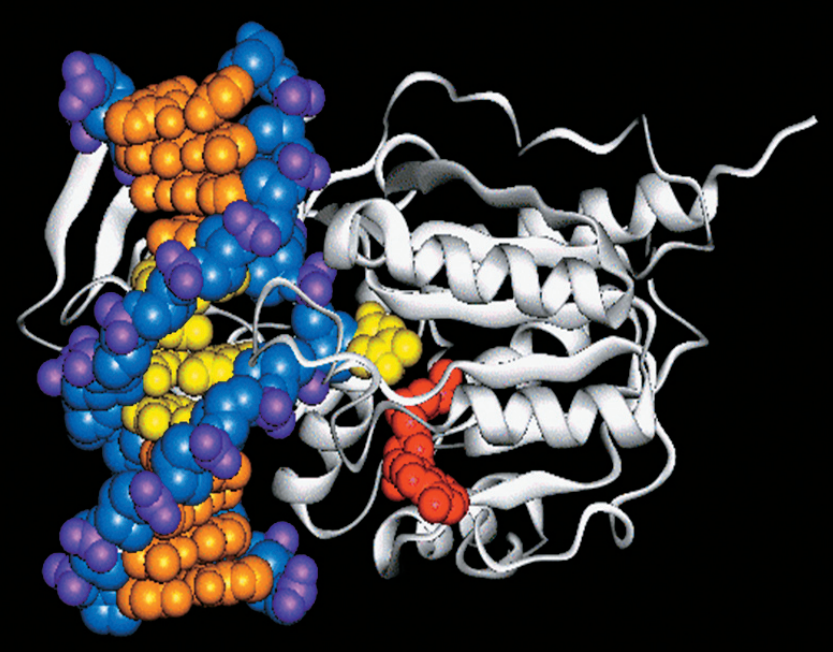
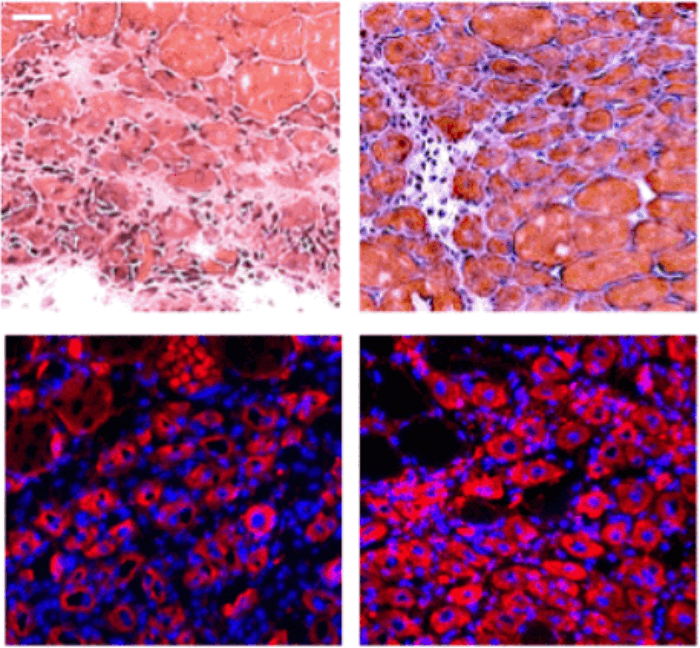
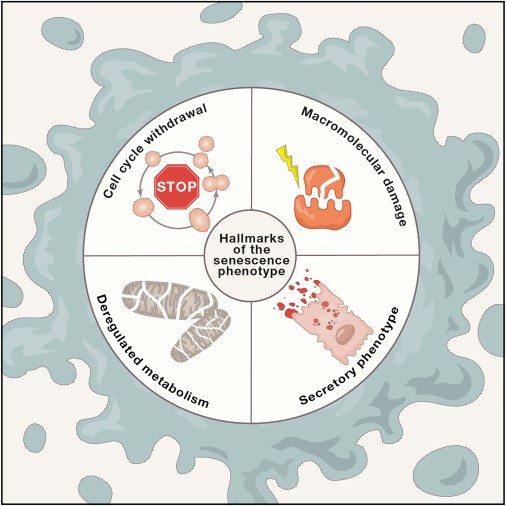
Рис. 2. Дифференцировка клеток нервной трубки мышиного эмбриона. Показаны поперечные срезы нервной трубки в районе будущего пояса передних конечностей (возраст эмбриона — 10,5 суток). На фотографиях клетки окрашены по-разному в зависимости от того, какие гены в них экспрессируются. На правой фотографии обведены границы доменов (область предшественников мотонейронов, pMN, — третья снизу, зеленые клетки снаружи от нее — нейроны, мигрирующие на периферию). NC — нотохорд, FP — донная пластинка, RP — потолочная пластинка. Длина масштабных линеек 50 мкм. Изображение из обсуждаемой статьи в Science
|
|
Разметка контролируется противоположными градиентами двух морфогенов: «вентрального» и «дорзального». На вентральной (нижней, брюшной) стороне нервной трубки, в так называемой донной пластинке (floor plate), а также в нотохорде, лежащем непосредственно под нервной трубкой, синтезируется белок Sonic hedgehog (Shh) — морфоген, участвующий в разметке многих частей организма. На дорзальной (спинной) стороне нервной трубки, в потолочной пластинке (roof plate), синтезируется другой многофункциональный морфоген, BMP (здесь, пожалуй, уместно лишний раз оговориться, что любое описание, тем более популярное, столь сложных биологических процессов — это всегда сильное упрощение, даже примитивизация: в действительности всё, как всегда, намного сложнее).
Таким образом, в нервной трубке создаются два противоположных градиента: сверху вниз растет концентрация Shh, снизу вверх нарастает воздействие BMP. В зависимости от сочетания этих концентраций прогениторные клетки нервной трубки (предшественники клеток нервной ткани) выбирают свою судьбу (cell fate): превращаются из потенциальных предшественников многих разных типов «взрослых» клеток в предшественников только одного типа клеток. Например, моторных или вставочных нейронов. Превращение состоит в том, что в клетках начинает экспрессироваться тот или иной набор транскрипционных факторов. В итоге нервная трубка подразделяется в дорзо-вентральном направлении на 13 доменов (зон), для каждого из которых характерна своя комбинация транскрипционных факторов. В частности, в третьем снизу домене, клетки которого будут в дальнейшем превращаться в мотонейроны спинного мозга, экспрессируется транскрипционный фактор Olig2, а шесть дорзальных доменов, которые дадут начало вставочным нейронам, можно опознать по экспрессии транскрипционного фактора Pax3.
Прогениторные клетки делятся, а потом в какой-то момент некоторые из них начинают превращаться (дифференцироваться) в нейроны того или иного типа. При этом прогениторные клетки остаются в центре, сохраняя контакт с невроцелем (спинномозговым каналом), и продолжают делиться, а нейроны мигрируют на периферию и больше не делятся.
Авторов интересовало, как меняются пропорции прогениторных доменов по мере роста эмбриона и каков механизм регуляции этих пропорций. Первый этап более или менее понятен: первичное подразделение на домены контролируется градиентами морфогенов, а вот что происходит дальше?
Для начала авторы убедились, что рост доменов — резко аллометрический, то есть их относительные размеры по мере роста нервной трубки сильно меняются. Например, домен pMN (область предшественников мотонейронов) сначала имеет довольно большой относительный размер, но потом, на девятый день развития эмбриона, он начинает быстро уменьшаться относительно других доменов (на рис. 2 справа он уже уменьшился). Авторы показали, что размер доменов в точности отражает число прогениторных клеток в них. Самое интересное, что относительные размеры доменов меняются похожим образом и у нормальных мышей, и у карликовых мышей-мутантов, и даже у куриного эмбриона. Значит, динамика пропорций доменов эволюционно консервативна — мало меняется в ходе эволюции. Из этого, в свою очередь, следует, что эта динамика и (или) те факторы, которые ее прямо или косвенно контролируют, очень важны для развивающегося организма.
Что же это за факторы? Авторы рассудили, что число прогениторных клеток в домене может меняться по четырем причинам: 1) пролиферация — деление клеток; 2) апоптоз — программируемая гибель клеток; 3) дифференцировка, то есть превращение прогениторной клетки в нейрон, который покидает домен, сокращая тем самым его размер; 4) смена клеточной идентичности, то есть превращение клетки одного домена в клетку другого домена (для этого должны отключиться одни транскрипционные факторы и включиться другие).
Используя большой арсенал современных методов генной инженерии, окраски и мечения клеток в зависимости от их состояния (наличия тех или иных белков, интенсивности синтеза ДНК и др.), авторы сумели оценить скорости всех четырех процессов на разных стадиях развития нервной трубки.
Скорость деления клеток во всех доменах оказалась практически одинаковой (кроме донной пластинки, где клетки делятся реже). По мере роста она немного снижается, причем это происходит синхронно во всех доменах. Значит, пропорции доменов определяются не темпом пролиферации.
Темп апоптоза оказался пренебрежимо низким по сравнению со скоростью деления клеток. Значит, апоптоз тоже не подходит на роль регулятора пропорций. Он активнее идет у карликовых мышей-мутантов — поэтому спинной мозг у них получается тоньше, но пропорции доменов и в этом случае определяются чем-то другим.
Скорость дифференцировки оказалась разной у разных доменов, и по мере роста она меняется тоже по-разному. Предшественники мотонейронов начинают дифференцироваться раньше других. Максимальная скорость дифференцировки у них тоже достигается раньше (60 hph, hours post headfold stage — через 60 часов после стадии головной складки, у мыши это 10 суток с начала развития). В это же самое время относительный размер домена pMN начинает быстро уменьшаться. В дорзальных доменах темп дифференцировки достигает максимума на 40 часов позже (100 hph).
Дальнейшие тесты и расчеты подтвердили, что различия в скорости роста доменов (а значит, и динамика их пропорций) на поздних стадиях (начиная с 40 hph) объясняются разной скоростью дифференцировки, которая у каждого домена своя. Те домены, в которых дифференцировка идет быстрее, растут медленнее (их относительный размер уменьшается), и наоборот.
Что же касается ранних стадий (<40 hph), то в это время различия в скорости дифференцировки не могут играть решающей роли — просто потому, что дифференцировка еще толком не началась. Чтобы понять, что происходит в этот период, авторам пришлось измерить темпы четвертого из перечисленных выше процессов — смены клеточной идентичности. Для этого использовалась хитроумная методика, позволяющая подсчитать клетки, которые раньше производили определенный транскрипционный фактор, а потом перестали это делать. Были изготовлены генно-модифицированные мыши, у которых, после введения определенного вещества, те клетки, в которых экспрессируется транскрипционный фактор Olig2 (это, напомним, характерная особенность клеток домена pMN — предшественников мотонейронов), начинают производить зеленый флуоресцирующий белок. Если клетка потом перестает производить Olig2 (то есть превращается в клетку другого домена), она всё равно продолжает светиться зеленым. Наличие же в клетке белка Olig2 можно установить при помощи окраски флуоресцирующими антителами (см. Immunostaining). Таким образом, клетки pMN получат две метки одновременно, а бывшие клетки pMN — только одну.
Выяснилось, что темп смены клеточной идентичности — перехода клеток из одного домена в другой путем выключения одних транскрипционных факторов и включения других — высок на ранних стадиях (<40 hph) и резко снижается на поздних.
Как мы помним, включение транскрипционных факторов, маркирующих домены, контролируется градиентами морфогенов. Поэтому данный факт — прозрачный намек на то, что вплоть до 40 hph судьба доменов и их границы определяются морфогенами. В дальнейшем влияние морфогенов снижается, и в дело вступает другой механизм регуляции роста доменов, связанный с темпом дифференцировки.
Чтобы окончательно убедиться в этом, авторы провели ряд экспериментов, в которых они искусственно ослабляли работу сигнального каскада Shh во всех доменах одновременно. Это делалось путем введения ингибитора — циклопамина (вещество получило такое название, потому что, подавляя работу Shh, оно может приводить к циклопии: ведь Shh играет важную роль в том числе и в правильном подразделении лица на две симметричные половинки). Оказалось, что подавление Shh сильно влияет на скорость роста и положение границ доменов до 40 hph, а на более поздних стадиях его эффект резко снижается.
Все эти результаты (а также ряд других, о которых мы умолчали для краткости, включая опыты на эмбрионе цыпленка, показавшие, что у птиц и млекопитающих все эти процессы очень похожи) говорят о том, что регуляция роста доменов осуществляется в два этапа. На первом этапе ведущую роль играют градиенты морфогенов Shh и BMP. Затем чувствительность прогениторных клеток к морфогенам снижается, и рост доменов переходит на «автономный режим» (саморегуляцию): теперь скорость роста и пропорции доменов определяются характерными для каждого домена темпами дифференцировки. От чего зависят эти темпы — следующий важный вопрос, который предстоит разрешить в будущем.
Данное исследование — небольшой, но важный шаг к решению актуальнейшей проблемы биологии: к пониманию того, как генетическая информация, через посредничество сложных процессов самоорганизации в ходе онтогенеза, воплощается в фенотипе. За конкретными деталями конкретных процессов, разгадываемых учеными, постепенно проступают общие принципы. В данном случае показан переход от «общесистемного» управления к локальному, автономному. Первичная разметка идет под контролем системных сигналов — градиентов разлитых в межклеточном пространстве веществ, производимых некими организующими центрами. Но затем размеченные участки развивающегося органа переходят на самоуправление (хотя, конечно, речь не идет о полной независимости от окружающих тканей и внешних сигналов; на поздних стадиях развития нервной трубки рост доменов продолжает отчасти контролироваться такими сигналами — на это есть ряд прямых указаний в обсуждаемой статье).
От себя добавлю, что опыт компьютерного моделирования онтогенеза (см.: Процессы самоорганизации в онтогенезе многоклеточных: опыт имитационного моделирования) помогает понять логику такой двухфазной регуляции. Разметить эмбрион на части при помощи градиентов морфогенов — просто, удобно и эффективно. Но получить желаемый взрослый фенотип (а не что-нибудь бесформенное с торчащими во все стороны ручками-ножками), не отключив своевременно чувствительность клеток к морфогенам и не переведя отделы тела на «саморегуляцию», очень нелегко. По мере роста эмбриона организаторы неизбежно смещаются, расходятся в пространстве, градиенты «плывут», а вслед за ними плывут и границы отделов тела, и весь план строения. Динамически регулировать производство морфогенов и чувствительность клеток к ним, подлаживаясь к меняющимся пропорциям частей тела, — в принципе можно, но это очень непростая программистская задача. Поэтому, моделируя онтогенезы сложных многоклеточных структур, мы обычно используем именно такую двухфазную схему регуляции, которую обнаружили в развитии нервной трубки авторы обсуждаемой статьи.
Источник: Anna Kicheva, Tobias Bollenbach, Ana Ribeiro, Helena Pérez Valle, Robin Lovell-Badge, Vasso Episkopou, James Briscoe. Coordination of progenitor specification and growth in mouse and chick spinal cord // Science. 2014. V. 345 P. 1577.

 Суть вот в чем:
В настоящий момент Google готовит наиболее неопределенный и долгосрочный маневр. Компания планирует запустить Calico — новая компания, которая будет сосредоточена на вопросах здравоохранения и старения, в частности. Независимой компанией будет управлять Артур Левинсон, бывший генеральный директор биотехнологической компании Genentech, который также будет инвестором. Левинсон, который начал свою карьеру как ученый и имеет степень доктора в области биохимии, планирует остаться в своей нынешней роли в качестве председателя совета директоров Genentech и Apple (должность, которую он взял на себя после смерти Стива Джобса в 2011 году). Google готовится сделать серьезную попытку увеличить продолжительность жизни человека.
На Google+ генеральный директор Google Ларри Пейдж написал: «Хорошо… вы, вероятно, думаете — ничего себе! Это сильно отличается от того, что Google делает сегодня. И вы правы. Но, как мы объяснили в нашем первом письме к акционерам, есть огромный потенциал технологий для улучшения жизни людей. Так что не удивляйтесь, если мы инвестируем в проекты, которые кажутся странными или спекулятивными по сравнению с нашим существующим интернет-бизнесом. И, пожалуйста, помните, что новые инвестиции, как Calico, очень маленькие по сравнению с нашим основным бизнесом».
Итак, как Calico собирается добиться увеличения длительности нашей жизни, и, возможно, в конечном счете излечить смерть? Пейдж не опубликовал какой-то конкретный план. На Google+ он говорит: «Еще прошло слишком мало времени, чтобы чем-то поделиться. Конечно, когда Артуру будет, что сказать, он обязательно этим поделится». Это неординарный проект, но это то, что любит Пейдж. Он хочет решать масштабные проблемы. Вот почему Google разрабатывает самоуправляемые автомобили и воздушные шары, плавающие в воздухе с интернет-соединениями. Google, у которого деньги куры не клюют и безграничные амбиции, видит себя в качестве единственной компанией, готовой взять на себя большие риски, как эти. «Я не предлагаю потратить все наши деньги на спорные вещи, но мы должны тратить соразмерный объем с тем, что обычные компании тратят на исследования и разработки, и тратить их на вещи, которые немного более долгосрочные и немного более амбициозные, чем обычно», — говорит Ларри.
Вот полный текст
Суть вот в чем:
В настоящий момент Google готовит наиболее неопределенный и долгосрочный маневр. Компания планирует запустить Calico — новая компания, которая будет сосредоточена на вопросах здравоохранения и старения, в частности. Независимой компанией будет управлять Артур Левинсон, бывший генеральный директор биотехнологической компании Genentech, который также будет инвестором. Левинсон, который начал свою карьеру как ученый и имеет степень доктора в области биохимии, планирует остаться в своей нынешней роли в качестве председателя совета директоров Genentech и Apple (должность, которую он взял на себя после смерти Стива Джобса в 2011 году). Google готовится сделать серьезную попытку увеличить продолжительность жизни человека.
На Google+ генеральный директор Google Ларри Пейдж написал: «Хорошо… вы, вероятно, думаете — ничего себе! Это сильно отличается от того, что Google делает сегодня. И вы правы. Но, как мы объяснили в нашем первом письме к акционерам, есть огромный потенциал технологий для улучшения жизни людей. Так что не удивляйтесь, если мы инвестируем в проекты, которые кажутся странными или спекулятивными по сравнению с нашим существующим интернет-бизнесом. И, пожалуйста, помните, что новые инвестиции, как Calico, очень маленькие по сравнению с нашим основным бизнесом».
Итак, как Calico собирается добиться увеличения длительности нашей жизни, и, возможно, в конечном счете излечить смерть? Пейдж не опубликовал какой-то конкретный план. На Google+ он говорит: «Еще прошло слишком мало времени, чтобы чем-то поделиться. Конечно, когда Артуру будет, что сказать, он обязательно этим поделится». Это неординарный проект, но это то, что любит Пейдж. Он хочет решать масштабные проблемы. Вот почему Google разрабатывает самоуправляемые автомобили и воздушные шары, плавающие в воздухе с интернет-соединениями. Google, у которого деньги куры не клюют и безграничные амбиции, видит себя в качестве единственной компанией, готовой взять на себя большие риски, как эти. «Я не предлагаю потратить все наши деньги на спорные вещи, но мы должны тратить соразмерный объем с тем, что обычные компании тратят на исследования и разработки, и тратить их на вещи, которые немного более долгосрочные и немного более амбициозные, чем обычно», — говорит Ларри.
Вот полный текст